
Evolución convergente
La evolución convergente es la evolución independiente de características similares en especies de diferentes períodos o épocas en el tiempo. La evolución convergente crea estructuras análogas que tienen una forma o función similar pero que no estaban presentes en el último ancestro común de esos grupos. El término cladístico para el mismo fenómeno es homoplasia. La evolución recurrente del vuelo es un ejemplo clásico, ya que los insectos voladores, las aves, los pterosaurios y los murciélagos han desarrollado de forma independiente la capacidad útil de vuelo. Las características funcionalmente similares que han surgido a través de la evolución convergente son análogas, mientras que las características homólogasLas estructuras o rasgos tienen un origen común pero pueden tener funciones diferentes. Las alas de pájaro, murciélago y pterosaurio son estructuras análogas, pero sus extremidades anteriores son homólogas y comparten un estado ancestral a pesar de cumplir funciones diferentes.
Lo opuesto a la convergencia es la evolución divergente, donde las especies relacionadas desarrollan diferentes rasgos. La evolución convergente es similar a la evolución paralela, que ocurre cuando dos especies independientes evolucionan en la misma dirección y, por lo tanto, adquieren características similares de manera independiente; por ejemplo, las ranas deslizantes han evolucionado en paralelo a partir de múltiples tipos de ranas arborícolas.
Se conocen muchos casos de evolución convergente en las plantas, incluido el desarrollo repetido de la fotosíntesis C 4, la dispersión de semillas por frutos carnosos adaptados para ser consumidos por animales y la carnivoría.
Visión de conjunto
En morfología, los rasgos análogos surgen cuando diferentes especies viven de manera similar y/o en un entorno similar y, por lo tanto, se enfrentan a los mismos factores ambientales. Cuando se ocupan nichos ecológicos similares (es decir, una forma de vida distintiva), problemas similares pueden conducir a soluciones similares. El anatomista británico Richard Owen fue el primero en identificar la diferencia fundamental entre analogías y homologías.
En bioquímica, las restricciones físicas y químicas sobre los mecanismos han provocado que algunos arreglos de sitios activos, como la tríada catalítica, evolucionen de forma independiente en superfamilias de enzimas separadas.
En su libro de 1989 Wonderful Life, Stephen Jay Gould argumentó que si uno pudiera "rebobinar la cinta de la vida [y] se encontraran nuevamente las mismas condiciones, la evolución podría tomar un curso muy diferente". Simon Conway Morris cuestiona esta conclusión, argumentando que la convergencia es una fuerza dominante en la evolución, y dado que las mismas restricciones ambientales y físicas están en juego, la vida inevitablemente evolucionará hacia un plan corporal "óptimo" y, en algún momento, la evolución está limitada. tropezar con la inteligencia, un rasgo actualmente identificado con al menos primates, córvidos y cetáceos.
Distinciones
Cladística
En cladística, una homoplasia es un rasgo compartido por dos o más taxones por cualquier motivo que no sea que comparten un ancestro común. Los taxones que comparten ascendencia son parte del mismo clado; la cladística busca ordenarlos según su grado de parentesco para describir su filogenia. Los rasgos homoplásticos causados por la convergencia son, por lo tanto, desde el punto de vista de la cladística, factores de confusión que podrían conducir a un análisis incorrecto.
Atavismo
En algunos casos, es difícil saber si un rasgo se perdió y luego volvió a evolucionar de manera convergente, o si un gen simplemente se apagó y luego se volvió a habilitar. Tal rasgo resurgido se llama atavismo. Desde un punto de vista matemático, un gen no utilizado (selectivamente neutral) tiene una probabilidad cada vez menor de retener la funcionalidad potencial a lo largo del tiempo. La escala de tiempo de este proceso varía mucho en diferentes filogenias; en mamíferos y aves, existe una probabilidad razonable de permanecer en el genoma en un estado potencialmente funcional durante unos 6 millones de años.
Evolución paralela vs. convergente
Cuando dos especies son similares en un carácter particular, la evolución se define como paralela si los ancestros también fueron similares, y convergente si no lo fueron. Algunos científicos han argumentado que existe un continuo entre la evolución paralela y la convergente, mientras que otros sostienen que, a pesar de cierta superposición, todavía existen distinciones importantes entre las dos.
Cuando las formas ancestrales no están especificadas o son desconocidas, o el rango de rasgos considerados no está claramente especificado, la distinción entre evolución paralela y convergente se vuelve más subjetiva. Por ejemplo, el sorprendente ejemplo de formas placentarias y marsupiales similares es descrito por Richard Dawkins en The Blind Watchmaker como un caso de evolución convergente, porque los mamíferos en cada continente tenían una larga historia evolutiva antes de la extinción de los dinosaurios bajo la cual acumular información relevante. diferencias
A nivel molecular
Proteínas
Sitios activos de proteasa
La enzimología de las proteasas proporciona algunos de los ejemplos más claros de evolución convergente. Estos ejemplos reflejan las limitaciones químicas intrínsecas de las enzimas, lo que lleva a la evolución a converger en soluciones equivalentes de forma independiente y repetida.
Las proteasas de serina y cisteína utilizan diferentes grupos funcionales de aminoácidos (alcohol o tiol) como nucleófilo. Para activar ese nucleófilo, orientan un residuo ácido y uno básico en una tríada catalítica. Las limitaciones químicas y físicas de la catálisis enzimática han provocado que arreglos de tríadas idénticos evolucionen independientemente más de 20 veces en diferentes superfamilias de enzimas.
Las treonina proteasas utilizan el aminoácido treonina como nucleófilo catalítico. A diferencia de la cisteína y la serina, la treonina es un alcohol secundario (es decir, tiene un grupo metilo). El grupo metilo de la treonina restringe en gran medida las posibles orientaciones de la tríada y el sustrato, ya que el metilo choca con el esqueleto de la enzima o la base de histidina. En consecuencia, la mayoría de las proteasas de treonina utilizan una treonina N-terminal para evitar tales choques estéricos. Varias superfamilias de enzimas evolutivamente independientes con diferentes pliegues de proteínas utilizan el residuo N-terminal como nucleófilo. Esta similitud del sitio activo pero la diferencia del pliegue de la proteína indica que el sitio activo evolucionó de manera convergente en esas familias.
Insulina de caracol y pescado en cono
Conus geographus produce una forma distinta de insulina que es más similar a las secuencias de proteína de insulina de pescado que a la insulina de moluscos más estrechamente relacionados, lo que sugiere una evolución convergente.
Na,K -ATPasa y resistencia de insectos a esteroides cardiotónicos
Existen muchos ejemplos de evolución convergente en insectos en términos de desarrollo de resistencia a nivel molecular a las toxinas. Un ejemplo bien caracterizado es la evolución de la resistencia a los esteroides cardiotónicos (CTS) a través de sustituciones de aminoácidos en posiciones bien definidas de la subunidad α de la Na,K-ATPasa (ATPalfa). La variación en ATPalpha ha sido examinada en varias especies adaptadas a CTS que abarcan 6 órdenes de insectos. Entre 21 especies adaptadas a CTS, 58 (76%) de 76 sustituciones de aminoácidos en sitios implicados en la resistencia a CTS ocurren en paralelo en al menos dos linajes. 30 de estas sustituciones (40%) ocurren en solo dos sitios en la proteína (posiciones 111 y 122). Las especies adaptadas a CTS también han desarrollado duplicaciones neofuncionalizadas recurrentes de ATPalfa, con patrones de expresión específicos de tejido convergentes.
Ácidos nucleicos
La convergencia ocurre a nivel de ADN y las secuencias de aminoácidos producidas al traducir genes estructurales en proteínas. Los estudios han encontrado convergencia en las secuencias de aminoácidos en murciélagos ecolocadores y delfines; entre mamíferos marinos; entre pandas gigantes y rojos; y entre los tilacinos y los cánidos. También se ha detectado convergencia en un tipo de ADN no codificante, elementos reguladores en cis, como en sus tasas de evolución; esto podría indicar una selección positiva o una selección purificadora relajada.
En morfología animal
Planes corporales
Animales nadadores, incluidos peces como arenques, mamíferos marinos como delfines e ictiosaurios (del Mesozoico), todos convergieron en la misma forma aerodinámica. Una forma similar y adaptaciones de natación están incluso presentes en moluscos, como Phylliroe. La forma del cuerpo fusiforme (un tubo cónico en ambos extremos) adoptada por muchos animales acuáticos es una adaptación que les permite viajar a alta velocidad en un entorno de alta resistencia. Se encuentran formas corporales similares en las focas sin orejas y las focas con orejas: todavía tienen cuatro patas, pero están fuertemente modificadas para nadar.
La fauna marsupial de Australia y los mamíferos placentarios del Viejo Mundo tienen varias formas sorprendentemente similares, desarrolladas en dos clados, aislados entre sí. El cuerpo y especialmente la forma del cráneo del tilacino (tigre de Tasmania o lobo de Tasmania) convergieron con los de Canidae como el zorro rojo, Vulpes vulpes.
- Convergencia de mamíferos marsupiales y placentarios
 esqueleto de zorro rojo
esqueleto de zorro rojo Cráneos de tilacino (izquierda), lobo de madera (derecha)
Cráneos de tilacino (izquierda), lobo de madera (derecha) esqueleto de tilacino
esqueleto de tilacino
Ecolocalización
Como adaptación sensorial, la ecolocalización ha evolucionado por separado en cetáceos (delfines y ballenas) y murciélagos, pero a partir de las mismas mutaciones genéticas.
Ojos
Uno de los ejemplos más conocidos de evolución convergente es el ojo de cámara de los cefalópodos (como el calamar y el pulpo), los vertebrados (incluidos los mamíferos) y los cnidarios (como las medusas). Su último ancestro común tenía como mucho una simple mancha fotorreceptora, pero una serie de procesos llevaron al refinamiento progresivo de los ojos de cámara, con una marcada diferencia: el ojo del cefalópodo está "conectado" en la dirección opuesta, con vasos sanguíneos y nerviosos que entran desde la parte posterior de la retina, en lugar de la parte frontal como en los vertebrados. Como resultado, los cefalópodos carecen de un punto ciego.
Vuelo
Las aves y los murciélagos tienen extremidades homólogas porque ambos se derivan en última instancia de los tetrápodos terrestres, pero sus mecanismos de vuelo son solo análogos, por lo que sus alas son ejemplos de convergencia funcional. Los dos grupos tienen vuelo propulsado, evolucionado de forma independiente. Sus alas difieren sustancialmente en la construcción. El ala de murciélago es una membrana estirada a través de cuatro dedos extremadamente alargados y las piernas. El perfil aerodinámico del ala del ave está hecho de plumas, fuertemente unidas al antebrazo (el cúbito) y los huesos altamente fusionados de la muñeca y la mano (el carpometacarpo), con solo restos diminutos de dos dedos restantes, cada uno sujetando una sola pluma. Entonces, mientras que las alas de los murciélagos y las aves son funcionalmente convergentes, no son anatómicamente convergentes.Las aves y los murciélagos también comparten una alta concentración de cerebrósidos en la piel de sus alas. Esto mejora la flexibilidad de la piel, un rasgo útil para los animales voladores; otros mamíferos tienen una concentración mucho más baja. Los pterosaurios extintos desarrollaron alas de forma independiente a partir de sus patas delanteras y traseras, mientras que los insectos tienen alas que evolucionaron por separado a partir de diferentes órganos.
Las ardillas voladoras y los planeadores del azúcar son muy parecidos en sus diseños corporales, con alas planeadoras extendidas entre sus extremidades, pero las ardillas voladoras son mamíferos placentarios, mientras que los planeadores del azúcar son marsupiales, muy separados dentro del linaje de los mamíferos.
Las polillas halcón colibrí y los colibríes han desarrollado patrones similares de vuelo y alimentación.
Piezas bucales de insectos
Las piezas bucales de los insectos muestran muchos ejemplos de evolución convergente. Las piezas bucales de diferentes grupos de insectos consisten en un conjunto de órganos homólogos, especializados para la ingesta dietética de ese grupo de insectos. La evolución convergente de muchos grupos de insectos condujo desde las piezas bucales mordedoras-masticadoras originales a diferentes tipos de funciones derivadas más especializadas. Estos incluyen, por ejemplo, la probóscide de los insectos que visitan las flores, como las abejas y los escarabajos de las flores, o las piezas bucales que muerden y chupan de los insectos chupadores de sangre, como las pulgas y los mosquitos.
Pulgares oponibles
Los pulgares oponibles que permiten agarrar objetos se asocian con mayor frecuencia con primates, como humanos, monos, simios y lémures. Los pulgares oponibles también evolucionaron en los pandas gigantes, pero estos tienen una estructura completamente diferente, tienen seis dedos, incluido el pulgar, que se desarrolla a partir de un hueso de la muñeca completamente separado de otros dedos.
Primates
 |  |  |
 A pesar del aclaramiento similar del color de la piel después de mudarse fuera de África, diferentes genes estaban involucrados en los linajes europeos (izquierda) y asiáticos orientales (derecha). A pesar del aclaramiento similar del color de la piel después de mudarse fuera de África, diferentes genes estaban involucrados en los linajes europeos (izquierda) y asiáticos orientales (derecha). |
La evolución convergente en humanos incluye el color de ojos azules y el color de piel claro. Cuando los humanos emigraron de África, se trasladaron a latitudes más septentrionales con luz solar menos intensa. Fue beneficioso para ellos reducir la pigmentación de su piel. Parece seguro que hubo cierto aclaramiento del color de la piel antes de que los linajes de Europa y Asia oriental se separaran, ya que existen algunas diferencias genéticas que aclaran la piel que son comunes a ambos grupos. Sin embargo, después de que los linajes se separaron y se aislaron genéticamente, la piel de ambos grupos se aclaró más y ese aclaramiento adicional se debió a diferentes cambios genéticos.
| humanos | lémures | ||
|---|---|---|---|
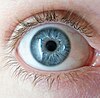 |  |  |  |
| A pesar de la similitud de apariencia, la base genética de los ojos azules es diferente en humanos y lémures. |
Los lémures y los humanos son ambos primates. Los primates ancestrales tenían ojos marrones, como la mayoría de los primates en la actualidad. La base genética de los ojos azules en humanos se ha estudiado en detalle y se sabe mucho al respecto. No es el caso de que un locus genético sea el responsable, por ejemplo, con el marrón dominante en el color de ojos azul. Sin embargo, un solo locus es responsable de alrededor del 80% de la variación. En los lémures, las diferencias entre los ojos azules y marrones no se conocen por completo, pero el mismo locus genético no está involucrado.
En las plantas
Fijacion de carbon
Si bien la evolución convergente a menudo se ilustra con ejemplos de animales, a menudo ha ocurrido en la evolución de las plantas. Por ejemplo, la fotosíntesis C 4, uno de los tres principales procesos bioquímicos de fijación de carbono, ha surgido de forma independiente hasta 40 veces. Alrededor de 7.600 especies de plantas de angiospermas utilizan la fijación de carbono C 4, con muchas monocotiledóneas que incluyen el 46% de los pastos como el maíz y la caña de azúcar, y dicotiledóneas que incluyen varias especies en Chenopodiaceae y Amaranthaceae.
Frutas
Un buen ejemplo de convergencia en plantas es la evolución de frutos comestibles como las manzanas. Estas pepitas incorporan (cinco) carpelos y sus tejidos accesorios formando el núcleo de la manzana, rodeados de estructuras externas al fruto botánico, el receptáculo o hipantio. Otras frutas comestibles incluyen otros tejidos vegetales; por ejemplo, la parte carnosa de un tomate son las paredes del pericarpio. Esto implica una evolución convergente bajo presión selectiva, en este caso la competencia por la dispersión de semillas por parte de los animales a través del consumo de frutos carnosos.
La dispersión de semillas por hormigas (myrmecochory) ha evolucionado independientemente más de 100 veces y está presente en más de 11,000 especies de plantas. Es uno de los ejemplos más dramáticos de evolución convergente en biología.
Carnívoro
El carnívoro ha evolucionado varias veces de forma independiente en plantas en grupos muy separados. En tres especies estudiadas, Cephalotus follicularis, Nepenthes alata y Sarracenia purpurea, ha habido convergencia a nivel molecular. Las plantas carnívoras secretan enzimas en el líquido digestivo que producen. Al estudiar las enzimas fosfatasa, glucósido hidrolasa, glucanasa, ARNasa y quitinasa, así como una proteína relacionada con la patogénesis y una proteína relacionada con la taumatina, los autores encontraron muchas sustituciones de aminoácidos convergentes. Estos cambios no estaban en los sitios catalíticos de las enzimas, sino en las superficies expuestas de las proteínas, donde podrían interactuar con otros componentes de la célula o el líquido digestivo. Los autores también encontraron que los genes homólogos en la planta no carnívora Arabidopsis thaliana tienden a aumentar su expresión cuando la planta está estresada, lo que lleva a los autores a sugerir que las proteínas que responden al estrés a menudo han sido cooptadas.en la evolución repetida de la carnivoría.
Métodos de inferencia
La reconstrucción filogenética y la reconstrucción del estado ancestral proceden asumiendo que la evolución ha ocurrido sin convergencia. Sin embargo, los patrones convergentes pueden aparecer en niveles más altos en una reconstrucción filogenética y, a veces, los investigadores los buscan explícitamente. Los métodos aplicados para inferir la evolución convergente dependen de si se espera una convergencia basada en patrones o basada en procesos. La convergencia basada en patrones es el término más amplio, cuando dos o más linajes desarrollan de forma independiente patrones de rasgos similares. La convergencia basada en procesos es cuando la convergencia se debe a fuerzas similares de selección natural.
Medidas basadas en patrones
Los métodos anteriores para medir la convergencia incorporan proporciones de distancia fenotípica y filogenética al simular la evolución con un modelo de movimiento browniano de evolución de rasgos a lo largo de una filogenia. Los métodos más recientes también cuantifican la fuerza de la convergencia. Un inconveniente a tener en cuenta es que estos métodos pueden confundir la estasis a largo plazo con la convergencia debido a las similitudes fenotípicas. La estasis ocurre cuando hay poco cambio evolutivo entre los taxones.
Las medidas basadas en la distancia evalúan el grado de similitud entre los linajes a lo largo del tiempo. Las medidas basadas en la frecuencia evalúan el número de linajes que han evolucionado en un espacio de rasgos particular.
Medidas basadas en procesos
Los métodos para inferir la convergencia basada en procesos ajustan los modelos de selección a una filogenia y datos de rasgos continuos para determinar si las mismas fuerzas selectivas han actuado sobre los linajes. Este utiliza el proceso de Ornstein-Uhlenbeck (OU) para probar diferentes escenarios de selección. Otros métodos se basan en una especificación a priori de dónde se han producido cambios en la selección.
Contenido relacionado
Nitrocelulosa
Desarrollo embrionario animal
Tablas de codones