
Quiasma óptico
En neuroanatomía, el quiasma óptico (del griego χίασμα 'cruce', del griego antiguo χιάζω 'marcar con una X'), es la parte del cerebro donde se cruzan los nervios ópticos. Se encuentra en la parte inferior del cerebro inmediatamente inferior al hipotálamo. El quiasma óptico se encuentra en todos los vertebrados, aunque en los ciclóstomos (lampreas y mixinos) se localiza dentro del cerebro.
Este artículo trata sobre el quiasma óptico de los vertebrados, que es el quiasma nervioso más conocido, pero no todos los quiasmas denotan un cruce de la línea media del cuerpo (p. ej., en algunos invertebrados, consulte Quiasma (anatomía)). Un cruce de nervios en la línea media dentro del cerebro se denomina decusación (ver Definición de tipos de cruces).
Estructura
Para los diferentes tipos de quiasma óptico, consulte
En todos los vertebrados, los nervios ópticos del ojo derecho e izquierdo se encuentran en la línea media del cuerpo, ventral al cerebro. En muchos vertebrados, el nervio óptico izquierdo cruza sobre el derecho sin fusionarse con él.
En vertebrados con una gran superposición de los campos visuales de los dos ojos, es decir, la mayoría de los mamíferos y aves, pero también anfibios, reptiles como los camaleones, los dos nervios ópticos se fusionan en el quiasma óptico. En tal quiasma óptico fusionado, parte de las fibras nerviosas no cruzan la línea media, sino que continúan hacia el tracto óptico del lado ipsilateral. Mediante esta decusación parcial, la parte del campo visual que está cubierta por ambos ojos se fusiona para permitir el procesamiento de la percepción de profundidad binocular por estereopsis (ver Figura 2).
En el caso de tal decusación parcial, las fibras del nervio óptico en los lados mediales de cada retina (que corresponden al lado lateral de cada hemicampo visual, porque la imagen está invertida) se cruzan hacia el lado opuesto de la línea media del cuerpo. La retina inferonasal está relacionada con la porción anterior del quiasma óptico, mientras que las fibras retinianas superonasales están relacionadas con la porción posterior del quiasma óptico.
El cruce parcial de las fibras del nervio óptico en el quiasma óptico permite que la corteza visual reciba el mismo campo visual hemisférico de ambos ojos. La superposición y el procesamiento de estas señales visuales monoculares permiten que la corteza visual genere visión binocular y estereoscópica. El resultado neto es que el hemisferio cerebral derecho procesa el hemicampo visual izquierdo y el hemisferio cerebral izquierdo procesa el hemicampo visual derecho.
Más allá del quiasma óptico, con fibras cruzadas y no cruzadas, los nervios ópticos se denominan vías ópticas. El tracto óptico se inserta en el techo óptico (en los mamíferos conocido como colículo superior) del mesencéfalo. En los mamíferos también se ramifican hacia el cuerpo geniculado lateral del tálamo, dándolos a su vez a la corteza occipital del cerebro.
Desarrollo en mamíferos
Durante el desarrollo, el cruce de los nervios ópticos está guiado principalmente por señales como netrina, hendidura, semaforina y efrina; y por morfógenos como sonic hedgehog (Shh) y Wnt. Esta navegación está mediada por el cono de crecimiento neuronal, una estructura que responde a las señales de los sistemas de señalización ligando-receptor que activan las vías aguas abajo que inducen cambios en el citoesqueleto. Los axones de las células ganglionares de la retina (RGC) que salen del ojo a través del nervio óptico están bloqueados para que no salgan de la vía de desarrollo mediante la inhibición de Slit2 y Sema5A, expresada bordeando la vía del nervio óptico. Ssh expresado en la línea media del sistema nervioso central inhibe el cruce antes del quiasma, donde se regula a la baja.La organización de los axones de RGC cambia de retinotópico a una orientación similar a una hoja plana a medida que se acercan al sitio del quiasma.
La mayoría de los axones de RGC cruzan la línea media en el diencéfalo ventral y continúan hasta el colículo superior contralateral. El número de axones que no cruzan la línea media y se proyectan ipsolateralmente depende del grado de visión binocular del animal (3% en ratones y 45% en humanos no cruzan). La efrina-B2 se expresa en la línea media del quiasma por la glía radial y actúa como una señal repulsiva para los axones que se originan en la retina ventrotemporal que expresa la proteína del receptor EphB1, dando lugar a la proyección ipsilateral o no cruzada. Los axones de RGC que se cruzan en el quiasma óptico están guiados por el factor de crecimiento endotelial vascular, VEGF-A, expresado en la línea media, que emite señales a través del receptor Neuropilin-1 (NRP1) expresado en los axones de RGC.El cruce del quiasma también es promovido por Nr-CAM (molécula de adhesión celular relacionada con Ng-CAM) y Semaphorin6D (Sema6D) expresado en la línea media, que forma un complejo que envía señales a los receptores Nr-CAM/Plexin-A1 al cruzar los axones RGC.
Otros animales
Mamíferos
Dado que todos los vertebrados, incluso los primeros fósiles y los modernos sin mandíbula, poseen un quiasma óptico, no se sabe cómo evolucionó. Se han propuesto varias teorías sobre la función del quiasma óptico en los vertebrados (ver teorías). Según la hipótesis del giro axial, el quiasma óptico se desarrolla como consecuencia de un giro en el embrión temprano.
En los gatos siameses con ciertos genotipos del gen albino, el cableado se interrumpe, con más nervios cruzados de lo normal. Dado que los gatos siameses, como los tigres albinos, también tienden a bizcar los ojos (estrabismo), se ha propuesto que este comportamiento podría compensar la cantidad anormal de decusación.
Cefalópodos e insectos
En cefalópodos e insectos, los tractos ópticos no cruzan la línea media del cuerpo, por lo que cada lado del cerebro procesa el ojo ipsilateral.
Historia
El cruce de fibras nerviosas y el impacto que esto tuvo en la visión probablemente fue identificado por primera vez por el médico persa "Esmail Jorjani", que parece ser Zayn al-Din Gorgani (1042-1137).
Imágenes Adicionales
 Esquema que muestra las conexiones centrales de los nervios ópticos y las vías ópticas.
Esquema que muestra las conexiones centrales de los nervios ópticos y las vías ópticas. Cerebro visto desde abajo, con el quiasma óptico visto en amarillo en el centro.
Cerebro visto desde abajo, con el quiasma óptico visto en amarillo en el centro. Transformaciones del campo visual hacia el mapa visual en la corteza visual primaria.
Transformaciones del campo visual hacia el mapa visual en la corteza visual primaria.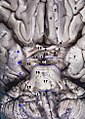 Cerebro y tronco encefálico vistos desde abajo
Cerebro y tronco encefálico vistos desde abajo Hemisferio izquierdo del cerebro visto en un espécimen cadavérico desde un lado, con el quiasma óptico marcado.
Hemisferio izquierdo del cerebro visto en un espécimen cadavérico desde un lado, con el quiasma óptico marcado. Cerebro, vista inferior, disección profunda.
Cerebro, vista inferior, disección profunda. Orientación del cruce y no cruce de axones durante el desarrollo.
Orientación del cruce y no cruce de axones durante el desarrollo.
Contenido relacionado
Glándula exocrina
Células somatótropas
Sistema nervioso parasimpático