
Médula espinal
La médula espinal es una estructura tubular larga y delgada formada por tejido nervioso, que se extiende desde el bulbo raquídeo en el tronco del encéfalo hasta la región lumbar de la columna vertebral. Encierra el canal central de la médula espinal, que contiene líquido cefalorraquídeo. El cerebro y la médula espinal juntos forman el sistema nervioso central (SNC). En los seres humanos, la médula espinal comienza en el hueso occipital, pasa por el agujero magno y entra en el canal espinal al comienzo de las vértebras cervicales. La médula espinal se extiende hasta entre la primera y la segunda vértebra lumbar, donde termina. La columna vertebral ósea envolvente protege la médula espinal relativamente más corta. Mide alrededor de 45 cm (18 pulgadas) de largo en hombres adultos y alrededor de 43 cm (17 pulgadas) de largo en mujeres adultas. El diámetro de la médula espinal varía de13 mm ( 1 ⁄ 2 in) en las regiones cervical y lumbar a 6,4 mm ( 1 ⁄ 4 in) en el área torácica.
La médula espinal funciona principalmente en la transmisión de señales nerviosas desde la corteza motora al cuerpo, y desde las fibras aferentes de las neuronas sensoriales a la corteza sensorial. También es un centro para coordinar muchos reflejos y contiene arcos reflejos que pueden controlar los reflejos de forma independiente. También es la ubicación de grupos de interneuronas espinales que forman los circuitos neuronales conocidos como generadores de patrones centrales. Estos circuitos se encargan de controlar las instrucciones motoras para movimientos rítmicos como caminar.
Estructura
La médula espinal es la vía principal de información que conecta el cerebro y el sistema nervioso periférico. Mucho más corta que su columna vertebral protectora, la médula espinal humana se origina en el tronco encefálico, pasa a través del agujero magno y continúa hasta el cono medular cerca de la segunda vértebra lumbar antes de terminar en una extensión fibrosa conocida como filum terminale.
Mide unos 45 cm (18 pulgadas) de largo en los hombres y unos 43 cm (17 pulgadas) en las mujeres, tiene forma ovoide y está agrandado en las regiones cervical y lumbar. El agrandamiento cervical, que se extiende desde la vértebra C5 hasta la T1, es de donde proviene la entrada sensorial y la salida motora va a los brazos y el tronco. El agrandamiento lumbar, ubicado entre L1 y S3, maneja la entrada sensorial y la salida motora que viene y va a las piernas.
La médula espinal se continúa con la porción caudal del bulbo raquídeo, desde la base del cráneo hasta el cuerpo de la primera vértebra lumbar. No recorre toda la longitud de la columna vertebral en adultos. Está hecho de 31 segmentos de los cuales se ramifican un par de raíces nerviosas sensoriales y un par de raíces nerviosas motoras. Las raíces nerviosas luego se fusionan en pares bilateralmente simétricos de nervios espinales. El sistema nervioso periférico está formado por estas raíces espinales, nervios y ganglios.
Las raíces dorsales son fascículos aferentes que reciben información sensorial de la piel, los músculos y los órganos viscerales para transmitirla al cerebro. Las raíces terminan en los ganglios de la raíz dorsal, que están compuestos por los cuerpos celulares de las neuronas correspondientes. Las raíces ventrales consisten en fibras eferentes que surgen de las neuronas motoras cuyos cuerpos celulares se encuentran en los cuernos grises ventrales (o anteriores) de la médula espinal.
La médula espinal (y el cerebro) están protegidos por tres capas de tejido o membranas llamadas meninges, que rodean el canal. La duramadre es la capa más externa y forma una capa protectora resistente. Entre la duramadre y el hueso circundante de las vértebras hay un espacio llamado espacio epidural. El espacio epidural está lleno de tejido adiposo y contiene una red de vasos sanguíneos. La aracnoides, la capa protectora intermedia, recibe su nombre por su apariencia abierta similar a una telaraña. El espacio entre la aracnoides y la piamadre subyacente se denomina espacio subaracnoideo. El espacio subaracnoideo contiene líquido cefalorraquídeo (LCR), que se puede extraer con una punción lumbar o un procedimiento de "punción espinal". La delicada piamadre, la capa protectora más interna, está estrechamente asociada con la superficie de la médula espinal. El cordón está estabilizado dentro de la duramadre por los ligamentos dentados de conexión, que se extienden desde la piamadre envolvente lateralmente entre las raíces dorsal y ventral. El saco dural termina en el nivel vertebral de la segunda vértebra sacra.
En corte transversal, la región periférica del cordón contiene tractos de sustancia blanca neuronal que contienen axones sensoriales y motores. En el interior de esta región periférica se encuentra la materia gris, que contiene los cuerpos de las células nerviosas dispuestas en las tres columnas grises que dan a la región su forma de mariposa. Esta región central rodea el canal central, que es una extensión del cuarto ventrículo y contiene líquido cefalorraquídeo.
La médula espinal tiene una sección transversal elíptica y está comprimida dorsolateralmente. Dos surcos prominentes, o surcos, corren a lo largo de su longitud. El surco mediano posterior es el surco en el lado dorsal, y la fisura mediana anterior es el surco en el lado ventral.
Segmentos
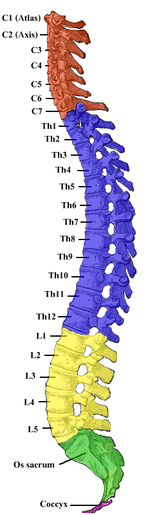
La médula espinal humana se divide en segmentos donde se forman pares de nervios espinales (mixtos; sensoriales y motores). Seis a ocho raicillas nerviosas motoras se ramifican de los surcos ventrales laterales derecho e izquierdo de una manera muy ordenada. Las raicillas nerviosas se combinan para formar raíces nerviosas. Del mismo modo, las raicillas nerviosas sensoriales se forman a partir de los surcos laterales dorsales derecho e izquierdo y forman raíces nerviosas sensoriales. Las raíces ventral (motora) y dorsal (sensorial) se combinan para formar nervios espinales (mixtos; motores y sensoriales), uno a cada lado de la médula espinal. Los nervios espinales, con la excepción de C1 y C2, se forman dentro del agujero intervertebral (FIV). Estas raicillas forman la demarcación entre los sistemas nerviosos central y periférico.
. Generalmente, los segmentos de la médula espinal no se corresponden con los niveles de las vértebras óseas. Como la médula espinal termina en el nivel L1-L2, otros segmentos de la médula espinal se ubicarían por encima del cuerpo de vértebras óseas correspondiente. Por ejemplo, el segmento espinal T11 está ubicado sobre las vértebras óseas T11, el segmento de la médula espinal sacra está ubicado sobre el cuerpo de las vértebras L1.
La columna gris, (como tres regiones de columnas grises) en el centro de la médula, tiene forma de mariposa y consta de cuerpos celulares de interneuronas, neuronas motoras, células de neuroglia y axones no mielinizados. La columna gris anterior y posterior se presentan como proyecciones de la sustancia gris y también se conocen como los cuernos de la médula espinal. Juntas, las columnas grises y la comisura gris forman la "H gris".
La sustancia blanca se encuentra fuera de la sustancia gris y consiste casi en su totalidad en axones motores y sensoriales mielinizados. Las "columnas" de sustancia blanca transportan información hacia arriba o hacia abajo de la médula espinal.
La médula espinal propiamente dicha termina en una región llamada conus medullaris, mientras que la piamadre continúa como una extensión llamada filum terminale, que ancla la médula espinal al cóccix. La cauda equina ("cola de caballo") es una colección de nervios inferiores al conus medullaris que continúan viajando a través de la columna vertebral hasta el cóccix. La cauda equina se forma porque la médula espinal deja de crecer en longitud alrededor de los cuatro años, aunque la columna vertebral continúa alargándose hasta la edad adulta. Esto da como resultado que los nervios espinales sacros se originen en la región lumbar superior. Por esa razón, la médula espinal ocupa solo dos tercios del canal vertebral. La parte inferior del canal vertebral está llena de líquido cefalorraquídeo (LCR) y el espacio se denomina cisterna lumbar.
Dentro del Sistema Nervioso Central (SNC), los cuerpos de las células nerviosas generalmente se organizan en grupos funcionales, llamados núcleos. Los axones dentro del SNC se agrupan en tractos.
Hay 31 segmentos de nervios de la médula espinal en una médula espinal humana:
- 8 segmentos cervicales que forman 8 pares de nervios cervicales (los nervios espinales C1 salen de la columna vertebral entre el agujero magno y la vértebra C1; los nervios C2 salen entre el arco posterior de la vértebra C1 y la lámina de C2; los nervios espinales C3-C8 pasan a través la FIV por encima de sus vértebras cervicales correspondientes, a excepción del par C8 que sale entre las vértebras C7 y T1)
- 12 segmentos torácicos que forman 12 pares de nervios torácicos
- 5 segmentos lumbares que forman 5 pares de nervios lumbares
- 5 segmentos sacros que forman 5 pares de nervios sacros
- 1 segmento coccígeo
| Especies | Cervical | torácico | Lumbar | Sacro | Caudal/Coccígea | Total |
|---|---|---|---|---|---|---|
| Perro | 8 | 13 | 7 | 3 | 5 | 36 |
| Gato | 8 | 13 | 7 | 3 | 5 | 36 |
| Vaca | 8 | 13 | 6 | 5 | 5 | 37 |
| Caballo | 8 | 18 | 6 | 5 | 5 | 42 |
| Cerdo | 8 | 15/14 | 6/7 | 4 | 5 | 38 |
| Humano | 8 | 12 | 5 | 5 | 1 | 31 |
| Ratón | 8 | 13 | 6 | 4 | 3 | 35 |
En el feto, los segmentos vertebrales se corresponden con los segmentos de la médula espinal. Sin embargo, debido a que la columna vertebral crece más que la médula espinal, los segmentos de la médula espinal no se corresponden con los segmentos vertebrales del adulto, particularmente en la parte inferior de la médula espinal. Por ejemplo, los segmentos de la médula espinal lumbar y sacra se encuentran entre los niveles vertebrales T9 y L2, y la médula espinal termina alrededor del nivel vertebral L1/L2, formando una estructura conocida como conus medullaris.
Aunque los cuerpos celulares de la médula espinal terminan alrededor del nivel vertebral L1/L2, los nervios espinales de cada segmento salen al nivel de la vértebra correspondiente. Para los nervios de la parte inferior de la médula espinal, esto significa que salen de la columna vertebral mucho más abajo (más caudalmente) que sus raíces. A medida que estos nervios viajan desde sus respectivas raíces hasta su punto de salida de la columna vertebral, los nervios de los segmentos espinales inferiores forman un haz llamado cauda equina.
Hay dos regiones donde la médula espinal se agranda:
- Agrandamiento cervical: corresponde aproximadamente a los nervios del plexo braquial, que inervan la extremidad superior. Incluye segmentos de la médula espinal desde aproximadamente C4 hasta T1. Los niveles vertebrales del agrandamiento son aproximadamente los mismos (C4 a T1).
- Agrandamiento lumbar: corresponde a los nervios del plexo lumbosacro, que inervan el miembro inferior. Comprende los segmentos de la médula espinal de L2 a S3 y se encuentra sobre los niveles vertebrales de T9 a T12.
Desarrollo
La médula espinal está formada por parte del tubo neural durante el desarrollo. Hay cuatro etapas de la médula espinal que surge del tubo neural: la placa neural, el pliegue neural, el tubo neural y la médula espinal. La diferenciación neural ocurre dentro de la porción de la médula espinal del tubo. A medida que el tubo neural comienza a desarrollarse, la notocorda comienza a secretar un factor conocido como Sonic hedgehog o SHH. Como resultado, la placa del suelo también comienza a secretar SHH, y esto inducirá a la placa basal a desarrollar neuronas motoras. Durante la maduración del tubo neural, sus paredes laterales se engrosan y forman un surco longitudinal llamado surco limitante. Esto extiende la longitud de la médula espinal en porciones dorsal y ventral también.Mientras tanto, el ectodermo suprayacente secreta proteína morfogenética ósea (BMP). Esto induce a la placa del techo a comenzar a secretar BMP, lo que inducirá a la placa alar a desarrollar neuronas sensoriales. Gradientes opuestos de morfógenos como BMP y SHH forman diferentes dominios de células en división a lo largo del eje ventral dorsal. Las neuronas del ganglio de la raíz dorsal se diferencian de los progenitores de la cresta neural. A medida que proliferan las células de las columnas dorsal y ventral, la luz del tubo neural se estrecha para formar el pequeño canal central de la médula espinal. La placa alar y la placa basal están separadas por el surco limitante. Además, la placa del suelo también segrega netrinas. Las netrinas actúan como quimioatrayentes para la decusación de las neuronas sensoriales del dolor y la temperatura en la placa alar a través de la comisura blanca anterior, donde luego ascienden hacia el tálamo. Tras el cierre del neuroporo caudal y la formación de los ventrículos del cerebro que contienen el tejido del plexo coroideo, el canal central de la médula espinal caudal se llena de líquido cefalorraquídeo.
Hallazgos anteriores de Viktor Hamburger y Rita Levi-Montalcini en embrión de pollo han sido confirmados por estudios más recientes que han demostrado que la eliminación de células neuronales por muerte celular programada (PCD) es necesaria para el correcto ensamblaje del sistema nervioso.
En general, se ha demostrado que la actividad embrionaria espontánea desempeña un papel en el desarrollo de las neuronas y los músculos, pero probablemente no esté involucrada en la formación inicial de conexiones entre las neuronas espinales.
Suministro de sangre
La médula espinal recibe sangre de tres arterias que recorren toda su longitud desde el cerebro y de muchas arterias que se acercan a ella a través de los lados de la columna vertebral. Las tres arterias longitudinales son la arteria espinal anterior y las arterias espinales posteriores derecha e izquierda. Estos viajan en el espacio subaracnoideo y envían ramas a la médula espinal. Forman anastomosis (conexiones) a través de las arterias medulares segmentarias anterior y posterior, que entran en la médula espinal en varios puntos a lo largo de su longitud. El flujo sanguíneo caudal real a través de estas arterias, derivado de la circulación cerebral posterior, es inadecuado para mantener la médula espinal más allá de los segmentos cervicales.
La principal contribución al suministro de sangre arterial de la médula espinal por debajo de la región cervical proviene de las arterias radiculares anterior y posterior dispuestas radialmente, que discurren hacia la médula espinal junto con las raíces nerviosas dorsal y ventral, pero con una excepción no se conectan directamente con cualquiera de las tres arterias longitudinales. Estas arterias radiculares intercostales y lumbares surgen de la aorta, proporcionan anastomosis importantes y complementan el flujo sanguíneo a la médula espinal. En los seres humanos, la mayor de las arterias radiculares anteriores se conoce como arteria de Adamkiewicz, o arteria radicular anterior magna (ARM), que generalmente surge entre L1 y L2, pero puede surgir en cualquier lugar entre T9 y L5.El flujo sanguíneo deteriorado a través de estas arterias radiculares críticas, especialmente durante procedimientos quirúrgicos que implican una interrupción abrupta del flujo sanguíneo a través de la aorta, por ejemplo, durante la reparación de un aneurisma aórtico, puede provocar un infarto de la médula espinal y paraplejia.
Función
Organización somatosensorial
En el tracto columna dorsal-lemniscus medial, el axón de una neurona primaria ingresa a la médula espinal y luego ingresa a la columna dorsal. Aquí la columna dorsal se conecta con el axón de la célula nerviosa. Si el axón primario entra por debajo del nivel espinal T6, el axón viaja en el fascículo grácil, la parte medial de la columna. Si el axón entra por encima del nivel T6, viaja en el fascículo cuneiforme, que es lateral al fascículo gracilis. De cualquier manera, el axón primario asciende a la parte inferior del bulbo raquídeo, donde deja su fascículo y hace sinapsis con una neurona secundaria en uno de los núcleos de la columna dorsal: ya sea el núcleo gracilis o el núcleo cuneatus, dependiendo de la vía que tomó. En este punto, el axón secundario sale de su núcleo y pasa anterior y medialmente. El conjunto de axones secundarios que hacen esto se conoce como fibras arqueadas internas. Las fibras arqueadas internas se decusan y continúan ascendiendo como el lemnisco medial contralateral. Los axones secundarios del lemnisco medial finalmente terminan en el núcleo posterolateral ventral (VPLN) del tálamo, donde hacen sinapsis con las neuronas terciarias. Desde allí, las neuronas terciarias ascienden por el brazo posterior de la cápsula interna y terminan en la corteza sensorial primaria.
La propiocepción de los miembros inferiores difiere de la de los miembros superiores y del tronco superior. Hay una vía de cuatro neuronas para la propiocepción de las extremidades inferiores. Esta vía sigue inicialmente la vía espinocerebelosa dorsal. Está organizado de la siguiente manera: receptores propioceptivos del miembro inferior → proceso periférico → ganglio de la raíz dorsal → proceso central → columna de Clarke → neurona de segundo orden → bulbo raquídeo (núcleo caudado) → neurona de tercer orden → VPLN del tálamo → neurona de cuarto orden → extremidad posterior de la cápsula interna → corona radiata → área sensorial del cerebro.
El sistema anterolateral funciona de manera algo diferente. Los axones de sus neuronas primarias ingresan a la médula espinal y luego ascienden uno o dos niveles antes de hacer sinapsis en la sustancia gelatinosa. El tracto que asciende antes de la sinapsis se conoce como tracto de Lissauer. Después de la sinapsis, los axones secundarios se decusan y ascienden en la porción lateral anterior de la médula espinal como el tracto espinotalámico. Este tracto asciende hasta el VPLN, donde hace sinapsis con las neuronas terciarias. Los axones neuronales terciarios luego viajan a la corteza sensorial primaria a través de la rama posterior de la cápsula interna.
Algunas de las "fibras del dolor" en la ALS se desvían de su camino hacia la VPLN. En una de esas desviaciones, los axones viajan hacia la formación reticular en el mesencéfalo. Luego, la formación reticular se proyecta a varios lugares, incluido el hipocampo (para crear recuerdos sobre el dolor), el núcleo centromediano (para causar un dolor difuso e inespecífico) y varias partes de la corteza. Además, algunos axones de ALS se proyectan hacia la sustancia gris periacueductal en la protuberancia, y los axones que forman la sustancia gris periacueductal luego se proyectan hacia el núcleo del rafe mayor, que se proyecta de regreso hacia donde proviene la señal de dolor y la inhibe. Esto ayuda a controlar la sensación de dolor hasta cierto punto.
Organización motora
| Nivel | Función motora |
|---|---|
| C1-C6 | Flexores de cuello |
| C1–T1 | extensores de cuello |
| C3, C4, C5 | Diafragma de suministro (principalmente C4) |
| C5, C6 | Mueva el hombro, levante el brazo (deltoides); codo flexible (bíceps) |
| C6 | rotar externamente (supinar) el brazo |
| C6, C7 | Extender codo y muñeca (tríceps y extensores de muñeca); muñeca pronada |
| C7, C8 | Muñeca flexible; suministrar pequeños músculos de la mano |
| T1–T6 | Intercostales y tronco arriba de la cintura |
| T7–L1 | Músculos abdominales |
| L1–L4 | Articulación de cadera flexible |
| L2, L3, L4 | Aducir muslo; Extender la pierna a la altura de la rodilla (cuádriceps femoral) |
| L4, L5, S1 | abducir el muslo; Flexionar la pierna a la altura de la rodilla (isquiotibiales); Dorsiflex pie (tibial anterior); Extender los dedos de los pies |
| L5, S1, S2 | Extienda la pierna a la altura de la cadera (glúteo mayor); flexionar el pie y flexionar los dedos de los pies |
El tracto corticoespinal sirve como vía motora para las señales neuronales motoras superiores que provienen de la corteza cerebral y de los núcleos motores primitivos del tronco del encéfalo.
Las neuronas motoras superiores corticales se originan en las áreas de Brodmann 1, 2, 3, 4 y 6 y luego descienden en el brazo posterior de la cápsula interna, a través de la crus cerebri, hacia abajo a través de la protuberancia y las pirámides medulares, donde aproximadamente el 90% de los axones cruzan al lado contralateral en la decusación de las pirámides. Luego descienden como el tracto corticoespinal lateral. Estos axones hacen sinapsis con las neuronas motoras inferiores en los cuernos ventrales de todos los niveles de la médula espinal. El 10% restante de los axones desciende en el lado ipsilateral como el tracto corticoespinal ventral. Estos axones también hacen sinapsis con las neuronas motoras inferiores en los cuernos ventrales. La mayoría de ellos cruzarán al lado contralateral del cordón (a través de la comisura blanca anterior) justo antes de hacer sinapsis.
Los núcleos del mesencéfalo incluyen cuatro tractos motores que envían axones de las neuronas motoras superiores a lo largo de la médula espinal hasta las neuronas motoras inferiores. Estos son el tracto rubroespinal, el tracto vestibuloespinal, el tracto tectoespinal y el tracto reticuloespinal. El tracto rubroespinal desciende con el tracto corticoespinal lateral y los tres restantes descienden con el tracto corticoespinal anterior.
La función de las neuronas motoras inferiores se puede dividir en dos grupos diferentes: el tracto corticoespinal lateral y el tracto espinal cortical anterior. El tracto lateral contiene axones de neuronas motoras superiores que hacen sinapsis con neuronas motoras inferiores dorsales laterales (DL). Las neuronas DL están implicadas en el control de las extremidades distales. Por lo tanto, estas neuronas DL se encuentran específicamente solo en los agrandamientos cervical y lumbosacro dentro de la médula espinal. No hay decusación en el tracto corticoespinal lateral después de la decusación en las pirámides medulares.
El tracto corticoespinal anterior desciende ipsilateralmente en la columna anterior, donde emergen los axones y hacen sinapsis con las neuronas motoras ventromediales inferiores (VM) en el asta ventral ipsilateralmente o se descusan en la comisura blanca anterior donde hacen sinapsis con las neuronas motoras inferiores VM contralateralmente. El tectoespinal, el vestibuloespinal y el reticuloespinal descienden ipsilateralmente en la columna anterior pero no hacen sinapsis a través de la comisura blanca anterior. Más bien, solo hacen sinapsis en las neuronas motoras inferiores de VM ipsilateralmente. Las neuronas motoras inferiores del VM controlan los grandes músculos posturales del esqueleto axial. Estas neuronas motoras inferiores, a diferencia de las del DL, están ubicadas en el asta ventral a lo largo de toda la médula espinal.
Tractos espinocerebelosos
La información propioceptiva en el cuerpo viaja por la médula espinal a través de tres vías. Por debajo de L2, la información propioceptiva viaja por la médula espinal en el tracto espinocerebeloso ventral. También conocido como el tracto espinocerebeloso anterior, los receptores sensoriales toman la información y viajan a la médula espinal. Los cuerpos celulares de estas neuronas primarias se encuentran en los ganglios de la raíz dorsal. En la médula espinal, los axones hacen sinapsis y los axones neuronales secundarios se decusan y luego viajan hasta el pedúnculo cerebeloso superior donde se decusan nuevamente. Desde aquí, la información se lleva a los núcleos profundos del cerebelo, incluidos los núcleos fastigiales e interpuestos.
Desde los niveles de L2 a T1, la información propioceptiva ingresa a la médula espinal y asciende ipsilateralmente, donde hace sinapsis en el núcleo de Clarke. Los axones neuronales secundarios continúan ascendiendo ipsilateralmente y luego pasan al cerebelo a través del pedúnculo cerebeloso inferior. Este tracto se conoce como el tracto espinocerebeloso dorsal.
Desde arriba de T1, los axones primarios propioceptivos ingresan a la médula espinal y ascienden ipsolateralmente hasta alcanzar el núcleo cuneiforme accesorio, donde hacen sinapsis. Los axones secundarios pasan al cerebelo a través del pedúnculo cerebeloso inferior donde, nuevamente, estos axones hacen sinapsis en los núcleos profundos del cerebelo. Este tracto se conoce como el tracto cuneocerebeloso.
La información motora viaja desde el cerebro por la médula espinal a través de los tractos descendentes de la médula espinal. Los tractos descendentes involucran dos neuronas: la neurona motora superior (UMN) y la neurona motora inferior (LMN). Una señal nerviosa viaja por la neurona motora superior hasta que hace sinapsis con la neurona motora inferior en la médula espinal. Luego, la neurona motora inferior conduce la señal nerviosa a la raíz espinal donde las fibras nerviosas eferentes llevan la señal motora hacia el músculo objetivo. Los tractos descendentes están compuestos de sustancia blanca. Hay varios tractos descendentes que cumplen diferentes funciones. Los tractos corticoespinales (lateral y anterior) son responsables de los movimientos coordinados de las extremidades.
Significación clínica
Un trastorno congénito es la diastematomielia en la que parte de la médula espinal se divide generalmente al nivel de las vértebras lumbares superiores. A veces, la división puede ser a lo largo de la médula espinal.
Lesión
Las lesiones de la médula espinal pueden ser causadas por un traumatismo en la columna vertebral (estiramiento, hematomas, aplicación de presión, corte, laceración, etc.). Los huesos vertebrales o los discos intervertebrales pueden romperse, lo que hace que la médula espinal sea perforada por un fragmento afilado de hueso. Por lo general, las víctimas de lesiones de la médula espinal sufrirán pérdida de sensibilidad en ciertas partes del cuerpo. En los casos más leves, la víctima solo puede sufrir la pérdida de la función de la mano o el pie. Las lesiones más graves pueden provocar paraplejía, tetraplejía (también conocida como cuadriplejía) o parálisis de todo el cuerpo debajo del sitio de la lesión en la médula espinal.
El daño a los axones de las neuronas motoras superiores en la médula espinal da como resultado un patrón característico de déficits ipsolaterales. Estos incluyen hiperreflexia, hipertonía y debilidad muscular. El daño de las neuronas motoras inferiores da como resultado su propio patrón característico de déficits. En lugar de un lado completo de los déficits, existe un patrón relacionado con el miotoma afectado por el daño. Además, las neuronas motoras inferiores se caracterizan por debilidad muscular, hipotonía, hiporreflexia y atrofia muscular.
El shock espinal y el shock neurogénico pueden ocurrir a partir de una lesión espinal. El shock espinal suele ser temporal, dura solo de 24 a 48 horas y es una ausencia temporal de las funciones sensoriales y motoras. El shock neurogénico dura semanas y puede conducir a una pérdida de tono muscular debido al desuso de los músculos debajo del sitio lesionado.
Las dos áreas de la médula espinal que se lesionan con mayor frecuencia son la columna cervical (C1-C7) y la columna lumbar (L1-L5). (La notación C1, C7, L1, L5 se refiere a la ubicación de una vértebra específica en la región cervical, torácica o lumbar de la columna). La lesión de la médula espinal también puede ser no traumática y estar causada por una enfermedad (mielitis transversa, poliomielitis, espina bífida, ataxia de Friedreich, tumor de la médula espinal, estenosis espinal, etc.)
A nivel mundial, se espera que haya alrededor de 40 a 80 casos de lesión de la médula espinal por cada millón de habitantes, y aproximadamente el 90 % de estos casos resultan de eventos traumáticos.
Las lesiones medulares reales o sospechadas requieren inmovilización inmediata, incluida la de la cabeza. Se necesitarán exploraciones para evaluar la lesión. Un esteroide, la metilprednisolona, puede ser de ayuda al igual que la fisioterapia y posiblemente los antioxidantes. Los tratamientos deben centrarse en limitar la muerte celular posterior a la lesión, promover la regeneración celular y reemplazar las células perdidas. La regeneración se facilita manteniendo la transmisión eléctrica en los elementos neurales.
Estenosis
La estenosis espinal en la región lumbar generalmente se debe a una hernia de disco, hipertrofia de la articulación facetaria y del ligamento amarillo, osteofito y espondilolistesis. La causa poco común de estenosis espinal lumbar se debe a la lipomatosis epidural espinal, una condición en la que hay un depósito excesivo de grasa en el espacio epidural, lo que provoca la compresión de la raíz nerviosa y la médula espinal. La grasa epidural se puede ver como de baja densidad en la tomografía computarizada y de alta intensidad en las imágenes de resonancia magnética de eco de espín rápido potenciadas en T2.
Tumores
Los tumores espinales pueden ocurrir en la médula espinal y pueden estar dentro (intradurales) o fuera (extradurales) de la duramadre.
Procedimientos
La médula espinal termina a nivel de las vértebras L1-L2, mientras que el espacio subaracnoideo —el compartimento que contiene el líquido cefalorraquídeo— se extiende hasta el borde inferior de S2. Las punciones lumbares en adultos se suelen realizar entre L3-L5 (nivel de cauda equina) para evitar daños en la médula espinal. En el feto, la médula espinal se extiende por toda la longitud de la columna y retrocede a medida que el cuerpo crece.
Imágenes Adicionales
 Anatomía seccional de la médula espinal. Animación en la referencia.
Anatomía seccional de la médula espinal. Animación en la referencia. Diagramas de la médula espinal.
Diagramas de la médula espinal. Sección transversal a través de la médula espinal a nivel torácico medio.
Sección transversal a través de la médula espinal a nivel torácico medio. Secciones transversales de la médula espinal en diferentes niveles.
Secciones transversales de la médula espinal en diferentes niveles.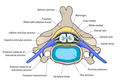 Vertebra cervical
Vertebra cervical Una porción de la médula espinal, mostrando su superficie lateral derecha. La duramadre se abre y se arregla para mostrar las raíces nerviosas.
Una porción de la médula espinal, mostrando su superficie lateral derecha. La duramadre se abre y se arregla para mostrar las raíces nerviosas. La médula espinal con duramadre cortada abierta, mostrando las salidas de los nervios espinales.
La médula espinal con duramadre cortada abierta, mostrando las salidas de los nervios espinales.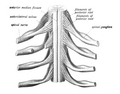 La médula espinal que muestra cómo las raíces anterior y posterior se unen en los nervios espinales.
La médula espinal que muestra cómo las raíces anterior y posterior se unen en los nervios espinales. La médula espinal que muestra cómo las raíces anterior y posterior se unen en los nervios espinales.
La médula espinal que muestra cómo las raíces anterior y posterior se unen en los nervios espinales. Una vista más larga de la médula espinal.
Una vista más larga de la médula espinal. Proyecciones de la médula espinal en los nervios (motor rojo, sensorial azul).
Proyecciones de la médula espinal en los nervios (motor rojo, sensorial azul). Proyecciones de la médula espinal en los nervios (motor rojo, sensorial azul).
Proyecciones de la médula espinal en los nervios (motor rojo, sensorial azul). Sección transversal de la médula espinal de conejo.
Sección transversal de la médula espinal de conejo. Corte transversal de médula espinal de rata adulta teñida con el método de Cajal.
Corte transversal de médula espinal de rata adulta teñida con el método de Cajal.
Contenido relacionado
Médula ósea
Dendrita
Glándula salival