
Hoja
Una hoja (hojas en plural) es el principal apéndice lateral del tallo de la planta vascular, generalmente por encima del suelo y especializado para la fotosíntesis. Las hojas, el tallo, la flor y el fruto juntos forman el sistema de brotes. Las hojas se denominan colectivamente follaje, como en "follaje de otoño". En la mayoría de las hojas, el tejido fotosintético primario es el mesófilo en empalizada y se encuentra en la parte superior del limbo o lámina de la hoja, pero en algunas especies, incluido el follaje maduro de Eucalyptus,el mesófilo en empalizada está presente en ambos lados y se dice que las hojas son isobilaterales. La mayoría de las hojas son aplanadas y tienen distintas superficies superior (adaxial) e inferior (abaxial) que difieren en color, vellosidad, número de estomas (poros que toman y expulsan gases), la cantidad y estructura de la cera epicuticular y otras características. Las hojas son en su mayoría de color verde debido a la presencia de un compuesto llamado clorofila que es esencial para la fotosíntesis ya que absorbe la energía de la luz del sol. Una hoja con parches o bordes de color más claro o blanco se llama hoja abigarrada.
Las hojas pueden tener muchas formas, tamaños y texturas diferentes. Las hojas anchas, planas y con nervadura compleja de las plantas con flores se conocen como megáfilas y las especies que las portan, en su mayoría, como plantas de hoja ancha o megáfilas, entre las que también se incluyen las acrogimnospermas y los helechos. En los licopodos, con diferentes orígenes evolutivos, las hojas son simples (con una sola nervadura) y se conocen como micrófilas.Algunas hojas, como las escamas de los bulbos, no están sobre el suelo. En muchas especies acuáticas, las hojas se sumergen en agua. Las plantas suculentas a menudo tienen hojas gruesas y jugosas, pero algunas hojas no tienen una función fotosintética importante y pueden estar muertas en la madurez, como en algunas catafilas y espinas. Además, varios tipos de estructuras similares a hojas que se encuentran en las plantas vasculares no son totalmente homólogas a ellas. Los ejemplos incluyen tallos de plantas aplanados llamados phylloclades y cladodes, y tallos de hojas aplanadas llamados phyllodes que difieren de las hojas tanto en su estructura como en su origen. Algunas estructuras de plantas no vasculares se ven y funcionan como hojas. Los ejemplos incluyen los fílidos de musgos y hepáticas.
Características generales
Las hojas son los órganos más importantes de la mayoría de las plantas vasculares.Las plantas verdes son autótrofas, lo que significa que no obtienen alimento de otros seres vivos, sino que crean su propio alimento mediante la fotosíntesis. Capturan la energía de la luz solar y la utilizan para producir azúcares simples, como glucosa y sacarosa, a partir de dióxido de carbono y agua. Luego, los azúcares se almacenan como almidón, luego se procesan mediante síntesis química en moléculas orgánicas más complejas, como proteínas o celulosa, el material estructural básico en las paredes celulares de las plantas, o se metabolizan mediante la respiración celular para proporcionar energía química para ejecutar procesos celulares. Las hojas extraen agua del suelo en la corriente de transpiración a través de un sistema de conducción vascular conocido como xilema y obtienen dióxido de carbono de la atmósfera por difusión a través de aberturas llamadas estomas en la capa exterior de la hoja (epidermis). mientras que las hojas están orientadas para maximizar su exposición a la luz solar. Una vez que se ha sintetizado el azúcar, debe transportarse a áreas de crecimiento activo, como los brotes y las raíces de las plantas. Las plantas vasculares transportan sacarosa en un tejido especial llamado floema. El floema y el xilema son paralelos entre sí, pero el transporte de materiales suele ser en direcciones opuestas. Dentro de la hoja, estos sistemas vasculares se ramifican para formar venas que suministran la mayor cantidad posible de hojas, asegurando que las células que realizan la fotosíntesis estén cerca del sistema de transporte. pero el transporte de materiales suele ser en direcciones opuestas. Dentro de la hoja, estos sistemas vasculares se ramifican para formar venas que suministran la mayor cantidad posible de hojas, asegurando que las células que realizan la fotosíntesis estén cerca del sistema de transporte. pero el transporte de materiales suele ser en direcciones opuestas. Dentro de la hoja, estos sistemas vasculares se ramifican para formar venas que suministran la mayor cantidad posible de hojas, asegurando que las células que realizan la fotosíntesis estén cerca del sistema de transporte.
Por lo general, las hojas son anchas, planas y delgadas (aplanadas dorsiventralmente), lo que maximiza el área de superficie directamente expuesta a la luz y permite que la luz penetre en los tejidos y llegue a los cloroplastos, promoviendo así la fotosíntesis. Están dispuestos en la planta para exponer sus superficies a la luz de la manera más eficiente posible sin sombrearse entre sí, pero hay muchas excepciones y complicaciones. Por ejemplo, las plantas adaptadas a condiciones de viento pueden tener hojas colgantes, como en muchos sauces y eucaliptos. La forma plana o laminar también maximiza el contacto térmico con el aire circundante, promoviendo el enfriamiento. Funcionalmente, además de realizar la fotosíntesis, la hoja es el sitio principal de transpiración, proporcionando la energía necesaria para extraer el flujo de transpiración desde las raíces y la gutación.
Muchas coníferas tienen hojas delgadas en forma de aguja o escamas que pueden ser ventajosas en climas fríos con frecuentes nevadas y heladas. Estos se interpretan como reducidos a partir de hojas megáfilas de sus antepasados del Devónico. Algunas formas de hojas están adaptadas para modular la cantidad de luz que absorben para evitar o mitigar el calor excesivo, el daño ultravioleta o la desecación, o para sacrificar la eficiencia de absorción de luz a favor de la protección contra la herbivoría. Para las xerófitas, la principal limitación no es el flujo o la intensidad de la luz, sino la sequía. Algunas plantas de ventana como las especies de Fenestraria y algunas especies de Haworthia como Haworthia tesselata y Haworthia truncata son ejemplos de xerófitas. yBulbine mesembryanthemoides.
Las hojas también funcionan para almacenar energía química y agua (especialmente en las suculentas) y pueden convertirse en órganos especializados que cumplen otras funciones, como los zarcillos de los guisantes y otras legumbres, las espinas protectoras de los cactus y las trampas para insectos en plantas carnívoras como Nepenthes y Sarracenia. Las hojas son las unidades estructurales fundamentales a partir de las cuales se construyen los conos en las gimnospermas (cada escama de cono es una hoja megafila modificada conocida como esporofila) y a partir de las cuales se construyen las flores en las plantas con flores.
La organización interna de la mayoría de los tipos de hojas ha evolucionado para maximizar la exposición de los orgánulos fotosintéticos, los cloroplastos, a la luz y aumentar la absorción de dióxido de carbono al mismo tiempo que se controla la pérdida de agua. Sus superficies están impermeabilizadas por la cutícula de la planta y el intercambio de gases entre las células mesófilas y la atmósfera está controlado por aberturas diminutas (longitud y anchura medidas en decenas de µm) llamadas estomas que se abren o cierran para regular la tasa de intercambio de dióxido de carbono, oxígeno, y vapor de agua dentro y fuera del sistema de espacio intercelular interno. La apertura estomática está controlada por la presión de turgencia en un par de células protectoras que rodean la apertura estomática. En cualquier centímetro cuadrado de la hoja de una planta, puede haber de 1.000 a 100.000 estomas.
La forma y la estructura de las hojas varían considerablemente de una especie a otra, dependiendo en gran medida de su adaptación al clima y la luz disponible, pero también de otros factores como el pastoreo de animales (como los ciervos), los nutrientes disponibles y la competencia ecológica de otras plantas.. También se producen cambios considerables en el tipo de hoja dentro de las especies, por ejemplo, a medida que una planta madura; como ejemplo, las especies de eucalipto comúnmente tienen hojas colgantes isobilaterales cuando maduran y dominan a sus vecinos; sin embargo, tales árboles tienden a tener hojas dorsiventrales erectas u horizontales como plántulas, cuando su crecimiento está limitado por la luz disponible.Otros factores incluyen la necesidad de equilibrar la pérdida de agua a alta temperatura y baja humedad frente a la necesidad de absorber el dióxido de carbono atmosférico. En la mayoría de las plantas, las hojas también son los órganos principales responsables de la transpiración y la gutación (perlas de líquido que se forman en los márgenes de las hojas).
Las hojas también pueden almacenar alimentos y agua, y se modifican en consecuencia para cumplir con estas funciones, por ejemplo, en las hojas de las plantas suculentas y en las escamas de los bulbos. La concentración de estructuras fotosintéticas en las hojas requiere que sean más ricas en proteínas, minerales y azúcares que, digamos, los tejidos del tallo leñoso. En consecuencia, las hojas ocupan un lugar destacado en la dieta de muchos animales.
En consecuencia, las hojas representan una gran inversión por parte de las plantas que las producen, y su retención o disposición son objeto de elaboradas estrategias para hacer frente a las presiones de las plagas, las condiciones estacionales y las medidas protectoras, como el crecimiento de espinas y la producción de fitolitos. ligninas, taninos y venenos.
Las plantas de hoja caduca en regiones templadas frías o frías generalmente pierden sus hojas en otoño, mientras que en áreas con una estación seca severa, algunas plantas pueden perder sus hojas hasta que finaliza la estación seca. En cualquier caso, se puede esperar que las hojas caídas aporten sus nutrientes retenidos al suelo donde caen.
En cambio, muchas otras plantas no estacionales, como las palmeras y las coníferas, conservan sus hojas durante largos periodos; Welwitschia conserva sus dos hojas principales a lo largo de una vida que puede superar los mil años.
Los órganos similares a hojas de las briófitas (p. ej., musgos y hepáticas), conocidos como fílidos, difieren mucho morfológicamente de las hojas de las plantas vasculares. En la mayoría de los casos, carecen de tejido vascular, tienen un grosor de una sola célula y no tienen cutícula, estomas o sistema interno de espacios intercelulares. (Los fílidos de la familia de los musgos Polytrichaceae son excepciones notables). Los fílidos de los briófitos solo están presentes en los gametofitos, mientras que, por el contrario, las hojas de las plantas vasculares solo están presentes en los esporofitos. Estos pueden desarrollarse aún más en estructuras vegetativas o reproductivas.
Hojas simples vascularizadas (micrófilas), como las del licopsido del Devónico temprano Baragwanathia, primero evolucionaron como enaciones, extensiones del tallo. Las hojas verdaderas o euphylls de mayor tamaño y con venación más compleja no se generalizaron en otros grupos hasta el período Devónico, momento en el que la concentración de dióxido de carbono en la atmósfera había disminuido significativamente. Esto ocurrió de forma independiente en varios linajes separados de plantas vasculares, en progimnospermas como Archaeopteris, en Sphenopsida, helechos y más tarde en las gimnospermas y angiospermas. Euphylls también se conoce como macrophylls o megaphylls (hojas grandes).
Morfología
Una hoja estructuralmente completa de una angiosperma consta de un pecíolo (tallo de la hoja), una lámina (lámina), estípulas (pequeñas estructuras ubicadas a ambos lados de la base del pecíolo) y una vaina. No todas las especies producen hojas con todos estos componentes estructurales. El tallo proximal o pecíolo se llama estípite en los helechos. La lámina es el componente plano y expandido de la hoja que contiene los cloroplastos. La vaina es una estructura, normalmente en la base, que abraza total o parcialmente el tallo por encima del nudo, donde se une este último. Las vainas de las hojas ocurren típicamente en pastos y Apiaceae (umbelíferas). Entre la vaina y la lámina, puede haber un pseudopeciolo, una estructura similar a un pecíolo. Los pseudopeciolos se encuentran en algunas monocotiledóneas, como plátanos, palmeras y bambúes.Las estípulas pueden ser conspicuas (p. ej., frijoles y rosas), caer pronto o no ser evidentes como en Moraceae o estar completamente ausentes como en Magnoliaceae. Un pecíolo puede estar ausente (apetiolado) o la lámina puede no ser laminar (aplanada). La enorme variedad que se muestra en la estructura de la hoja (anatomía) de una especie a otra se presenta en detalle a continuación en morfología. El pecíolo une mecánicamente la hoja a la planta y proporciona la ruta para la transferencia de agua y azúcares hacia y desde la hoja. La lámina es típicamente la ubicación de la mayoría de la fotosíntesis. El ángulo superior (adaxial) entre una hoja y un tallo se conoce como axila de la hoja. A menudo es la ubicación de un brote. Las estructuras ubicadas allí se denominan "axilares".
Las características externas de las hojas, como la forma, el margen, los pelos, el pecíolo y la presencia de estípulas y glándulas, son frecuentemente importantes para identificar plantas a nivel de familia, género o especie, y los botánicos han desarrollado una rica terminología para describir las características de las hojas. Las hojas casi siempre tienen un crecimiento determinado. Crecen a un patrón y forma específicos y luego se detienen. Otras partes de la planta, como los tallos o las raíces, tienen un crecimiento indeterminado y, por lo general, continuarán creciendo mientras tengan los recursos para hacerlo.
El tipo de hoja suele ser característico de una especie (monomórfica), aunque algunas especies producen más de un tipo de hoja (dimórfica o polimórfica). Las hojas más largas son las de la palmera rafia, R. regalis, que pueden medir hasta 25 m (82 pies) de largo y 3 m (9,8 pies) de ancho. La terminología asociada con la descripción de la morfología de la hoja se presenta, en forma ilustrada, en Wikilibros.
Cuando las hojas son basales y yacen en el suelo, se las denomina postradas.
Tipos básicos de hojas
Se dice que las plantas perennes cuyas hojas se desprenden anualmente tienen hojas caducas, mientras que las hojas que permanecen durante el invierno son las perennes. Las hojas unidas a los tallos por pedúnculos (conocidos como pecíolos) se llaman pecioladas, y si están unidas directamente al tallo sin pecíolo, se llaman sésiles.
- Los helechos tienen hojas.
- Las hojas de las coníferas suelen tener forma de aguja o punzón o escamas, por lo general son de hoja perenne, pero a veces pueden ser de hoja caduca. Por lo general, tienen una sola vena.
- Hojas de plantas con flores (angiospermas): la forma estándar incluye estípulas, un pecíolo y una lámina.
- Los licofitos tienen micrófilos.
- Las hojas de vaina son del tipo que se encuentra en la mayoría de los pastos y en muchas otras monocotiledóneas.
- Otras hojas especializadas incluyen las de Nepenthes, una planta de jarra.
Las hojas de las dicotiledóneas tienen láminas con vegetación pinnada (donde las venas principales divergen de una vena central grande y tienen redes de conexión más pequeñas entre ellas). Con menos frecuencia, las hojas de las dicotiledóneas pueden tener venación palmeada (varias venas grandes que divergen desde el pecíolo hasta los bordes de las hojas). Finalmente, algunos exhiben venación paralela.
Las hojas de monocotiledóneas en climas templados suelen tener hojas estrechas y, por lo general, nervaduras paralelas que convergen en las puntas o bordes de las hojas. Algunos también tienen venación pinnada.
Arreglo en el tallo
La disposición de las hojas sobre el tallo se conoce como filotaxis. En la naturaleza se dan una gran variedad de patrones filotácticos:AlternoUna hoja, rama o parte de la flor se adhiere a cada punto o nudo del tallo y las hojas alternan la dirección, en mayor o menor grado, a lo largo del tallo.BasalSurgido de la base del tallo.caulinaSurgido del tallo aéreo.OpuestoDos hojas, ramas o partes de flores se unen en cada punto o nudo del tallo. Los archivos adjuntos de hoja se emparejan en cada nodo.DecusadoUna disposición opuesta en la que cada par sucesivo gira 90° con respecto al anterior.Verticilado o verticiladoTres o más hojas, ramas o partes de flores se unen en cada punto o nudo del tallo. Al igual que con las hojas opuestas, los verticilos sucesivos pueden o no estar decusados, girados la mitad del ángulo entre las hojas en el verticilo (es decir, verticilos sucesivos de tres girados 60°, verticilos de cuatro girados 45°, etc.). Las hojas opuestas pueden aparecer verticiladas cerca de la punta del tallo. Pseudoverticillate describe un arreglo que solo parece verticilado, pero en realidad no lo es.rosuladoLas hojas forman una roseta.filasEl término dístico significa literalmente dos filas. Las hojas en este arreglo pueden ser alternas u opuestas en su unión. El término, 2-ranked, es equivalente. A veces se encuentran los términos tristichous y tetrastichous. Por ejemplo, las "hojas" (en realidad microfilas) de la mayoría de las especies de Selaginella son tetrásticas, pero no decusadas.
En los modelos matemáticos más simples de filotaxis, el vértice del tallo se representa como un círculo. Cada nuevo nodo se forma en el vértice y se gira en un ángulo constante con respecto al nodo anterior. Este ángulo se llama ángulo de divergencia. El número de hojas que crecen de un nudo depende de la especie de planta. Cuando una sola hoja crece de cada nudo, y cuando el tallo se mantiene recto, las hojas forman una hélice.
El ángulo de divergencia a menudo se representa como una fracción de una rotación completa alrededor del vástago. Una fracción de rotación de 1/2 (un ángulo de divergencia de 180°) produce un arreglo alternativo, como en Gasteria o el abanico-aloe Kumara plicatilis. Las fracciones de rotación de 1/3 (ángulos de divergencia de 120°) ocurren en haya y avellano. Roble y albaricoque rotan en 2/5, girasoles, álamos y perales en 3/8, y en sauces y almendros la fracción es de 5/13. Estos arreglos son periódicos. El denominador de la fracción de rotación indica el número de hojas en un período, mientras que el numerador indica el número de vueltas o giros completos realizados en un período. Por ejemplo:
- 180° (o 1 ⁄ 2 ): dos hojas en un círculo (hojas alternas)
- 120° (o 1 ⁄ 3 ): tres hojas en un círculo
- 144° (o 2 ⁄ 5 ): cinco hojas en dos giros
- 135° (o 3 ⁄ 8 ): ocho hojas en tres giros.
La mayoría de los ángulos de divergencia están relacionados con la secuencia de números de Fibonacci F n. Esta secuencia comienza 1, 1, 2, 3, 5, 8, 13; cada término es la suma de los dos anteriores. Las fracciones de rotación son a menudo cocientes F n / F n + 2 de un número de Fibonacci por el número dos términos más adelante en la secuencia. Este es el caso de las fracciones 1/2, 1/3, 2/5, 3/8 y 5/13. La relación entre números de Fibonacci sucesivos tiende a la proporción áurea φ = (1 + √5)//2. Cuando un círculo se divide en dos arcos cuyas longitudes están en la proporción 1:φ, el ángulo formado por el arco más pequeño es el ángulo áureo, que es 1/φ × 360° ≈ 137,5°. Debido a esto, muchos ángulos de divergencia son de aproximadamente 137,5°.
En las plantas donde crece un par de hojas opuestas de cada nudo, las hojas forman una doble hélice. Si los nodos no giran (una fracción de rotación de cero y un ángulo de divergencia de 0°), las dos hélices se convierten en un par de líneas paralelas, creando una disposición dística como en los arces o los olivos. Más común en un patrón decusado, en el que cada nodo gira 1/4 (90°) como en la hierba albahaca. Las hojas de plantas tricusadas como Nerium oleander forman una triple hélice.
Las hojas de algunas plantas no forman hélices. En algunas plantas, el ángulo de divergencia cambia a medida que la planta crece. En la filotaxis del orixato, llamada así por Orixa japonica, el ángulo de divergencia no es constante. En cambio, es periódica y sigue la secuencia 180°, 90°, 180°, 270°.
Divisiones de la hoja
Se pueden describir dos formas básicas de hojas considerando la forma en que se divide la hoja (lámina). Una hoja simple tiene una lámina no dividida. Sin embargo, la hoja se puede diseccionar para formar lóbulos, pero los espacios entre los lóbulos no llegan a la nervadura principal. Una hoja compuestatiene una hoja completamente subdividida, cada hojuela de la hoja está separada a lo largo de una vena principal o secundaria. Los folíolos pueden tener pecíolos y estípulas, los equivalentes de los pecíolos y estípulas de las hojas. Debido a que cada folleto puede parecer una hoja simple, es importante reconocer dónde se encuentra el pecíolo para identificar una hoja compuesta. Las hojas compuestas son una característica de algunas familias de plantas superiores, como las Fabaceae. La vena media de una hoja compuesta o una fronda, cuando está presente, se llama raquis.Compuesto palmeadoLas hojas tienen los folíolos que irradian desde el final del pecíolo, como los dedos de la palma de una mano; por ejemplo, Cannabis (cáñamo) y Aesculus (castaños).Compuesto pinnadoLas hojas tienen los folíolos dispuestos a lo largo del nervio principal o medio.impar pinnadaCon prospecto terminal; por ejemplo, Fraxinus (fresno).incluso pinnadasCareciendo de un prospecto terminal; por ejemplo, Swietenia (caoba). Un tipo específico de par pinnado es bipinnado, donde las hojas solo constan de dos folíolos; por ejemplo, Himenaea.Compuesto bipinnadoLas hojas se dividen dos veces: los folíolos se disponen a lo largo de una nervadura secundaria que es una de varias ramificaciones del raquis. Cada folleto se llama pinnule. El grupo de pinnules en cada vena secundaria forma un pinna ; por ejemplo, Albizia (árbol de la seda).Trifoliado (o trifoliolado)Una hoja pinnada con solo tres folíolos; por ejemplo, Trifolium (trébol), Laburnum (laburnum) y algunas especies de Toxicodendron (por ejemplo, hiedra venenosa).Tipos de superficiePinnadamente disecado hasta la vena central, pero con los folíolos no completamente separados; por ejemplo, Polypodium, algunos Sorbus (rayos blancos). En las hojas con venas pinnadas, la vena central se conoce como nervadura central.
Características del pecíolo
Se dice que las hojas que tienen un pecíolo (peciolo) son pecioladas.
Las hojas sésiles (epetioladas) no tienen pecíolo y la hoja se adhiere directamente al tallo. Las hojas subpecioladas son casi pecioladas o tienen un pecíolo extremadamente corto y pueden parecer sésiles.
En las hojas entrelazadas o decurrentes, el limbo rodea parcialmente al tallo.
Cuando la base de la hoja rodea completamente el tallo, se dice que las hojas están perfoliadas, como en Eupatorium perfoliatum.
En las hojas peltadas, el pecíolo se adhiere al limbo por dentro del margen del limbo.
En algunas especies de Acacia, como el árbol koa ( Acacia koa ), los pecíolos se expanden o ensanchan y funcionan como láminas de hojas; estos se llaman filodes. Puede haber o no hojas pinnadas normales en la punta del filodio.
Una estípula, presente en las hojas de muchas dicotiledóneas, es un apéndice a cada lado en la base del pecíolo, que se asemeja a una hoja pequeña. Las estípulas pueden ser duraderas y no desprenderse (una hoja estipulada, como en las rosas y los frijoles), o desprenderse a medida que la hoja se expande, dejando una cicatriz estipulada en la ramita (una hoja exestipulada). La situación, disposición y estructura de las estipulaciones se denomina "estipulación".libres, lateralesComo en Hibisco.adnatoFusionado a la base del pecíolo, como en Rosa.ocreatoProvisto de ocre, o estípulas en forma de vaina, como en Polygonaceae; por ejemplo, ruibarbo.Rodeando la base del pecíolointerpeciolarEntre los pecíolos de dos hojas opuestas, como en Rubiaceae.intrapeciolarEntre el pecíolo y el tallo subyacente, como en Malpighiaceae.
Venas
Las venas (a veces denominadas nervios) constituyen uno de los rasgos o características más visibles de las hojas. Las venas de una hoja representan la estructura vascular del órgano, se extienden hacia la hoja a través del pecíolo y proporcionan transporte de agua y nutrientes entre la hoja y el tallo, y juegan un papel crucial en el mantenimiento del estado hídrico de la hoja y la capacidad fotosintética. También juegan un papel en el soporte mecánico de la hoja. Dentro de la lámina de la hoja, mientras que algunas plantas vasculares poseen una sola vena, en la mayoría esta vasculatura generalmente se divide (ramifica) de acuerdo con una variedad de patrones (venación) y forma haces cilíndricos, generalmente ubicados en el plano medio del mesófilo. entre las dos capas de la epidermis.Este patrón es a menudo específico de los taxones, y de los cuales las angiospermas poseen dos tipos principales, paralelos y reticulados (como una red). En general, la nervadura paralela es típica de las monocotiledóneas, mientras que la reticulada es más típica de las eudicotiledóneas y las magnólidas ("dicotiledóneas"), aunque hay muchas excepciones.
La vena o venas que ingresan a la hoja desde el pecíolo se denominan venas primarias o de primer orden. Las venas que se ramifican de estos son venas secundarias o de segundo orden. Estas venas primarias y secundarias se consideran venas mayores o de menor orden, aunque algunos autores las incluyen de tercer orden. Cada ramificación subsiguiente está numerada secuencialmente, y estas son las venas de orden superior, estando asociada cada ramificación con un diámetro de vena más estrecho.En las hojas con nervaduras paralelas, las nervaduras primarias corren paralelas y equidistantes entre sí durante la mayor parte de la longitud de la hoja y luego convergen o se fusionan (anastomosan) hacia el ápice. Por lo general, muchas venas menores más pequeñas interconectan estas venas primarias, pero pueden terminar con terminaciones de venas muy finas en el mesófilo. Las venas menores son más típicas de las angiospermas, que pueden tener hasta cuatro órdenes superiores.En contraste, las hojas con nervadura reticulada tienen una nervadura primaria única (a veces más) en el centro de la hoja, denominada nervadura central o costa, y se continúa con la vasculatura del pecíolo más proximalmente. La nervadura central luego se ramifica en varias venas secundarias más pequeñas, también conocidas como venas de segundo orden, que se extienden hacia los márgenes de las hojas. Estos a menudo terminan en un hidátodo, un órgano secretor, en el margen. A su vez, las venas más pequeñas se ramifican a partir de las venas secundarias, conocidas como venas terciarias o de tercer orden (o superior), formando un denso patrón reticulado. Las áreas o islas de mesófilo que se encuentran entre las venas de orden superior se denominan areolas. Algunas de las venas más pequeñas (venillas) pueden terminar en las areolas, proceso conocido como areolación.Estas venas menores actúan como sitios de intercambio entre el mesófilo y el sistema vascular de la planta. Así, las nervaduras menores recogen los productos de la fotosíntesis (fotosintato) de las células donde se lleva a cabo, mientras que las nervaduras mayores se encargan de su transporte fuera de la hoja. Al mismo tiempo, el agua se transporta en la dirección opuesta.
El número de terminaciones de las venas es muy variable, al igual que si las venas de segundo orden terminan en el margen o se vinculan con otras venas. Hay muchas variaciones elaboradas en los patrones que forman las venas de las hojas, y éstas tienen implicaciones funcionales. De estos, las angiospermas tienen la mayor diversidad. Dentro de estos, las venas principales funcionan como red de soporte y distribución para las hojas y están correlacionadas con la forma de la hoja. Por ejemplo, la nervadura paralela que se encuentra en la mayoría de las monocotiledóneas se correlaciona con la forma de hoja alargada y la base ancha de la hoja, mientras que la nervadura reticulada se observa en hojas enteras simples, mientras que las hojas digitadas suelen tener una nervadura en la que tres o más nervaduras primarias divergen radialmente desde un solo punto..
En términos evolutivos, los taxones emergentes tempranos tienden a tener ramificaciones dicotómicas con sistemas reticulados que emergen más tarde. Las venas aparecieron en el período Pérmico (299-252 millones de años), antes de la aparición de las angiospermas en el Triásico (252-201 millones de años), durante el cual apareció la jerarquía de las venas que permitió una mayor función, un tamaño de hoja más grande y la adaptación a una variedad más amplia de condiciones climáticas.. Aunque es el patrón más complejo, las venas ramificadas parecen ser plesiomórficas y de alguna forma estaban presentes en plantas con semillas antiguas desde hace 250 millones de años. Una venación pseudo-reticulada que en realidad es una penniparalela altamente modificada es una autapomorfía de algunas Melanthiaceae, que son monocotiledóneas; por ejemplo, Paris quadrifolia(Nudo del verdadero amante). En hojas con venación reticulada, las venas forman una matriz de andamiaje que imparte rigidez mecánica a las hojas.
Cambios de morfología dentro de una sola planta
HomoblastiaCaracterística en la que una planta tiene pequeños cambios en el tamaño de las hojas, la forma y el hábito de crecimiento entre las etapas juvenil y adulta, en contraste con;heteroblastiaCaracterística en la que una planta tiene cambios marcados en el tamaño de la hoja, la forma y el hábito de crecimiento entre las etapas juvenil y adulta.
Anatomía
Características de mediana escala
Las hojas normalmente están muy vascularizadas y suelen tener redes de haces vasculares que contienen xilema, que suministra agua para la fotosíntesis, y floema, que transporta los azúcares producidos por la fotosíntesis. Muchas hojas están cubiertas de tricomas (pequeños pelos) que tienen diversas estructuras y funciones.

Características a pequeña escala
Los principales sistemas tisulares presentes son
- La epidermis, que cubre las superficies superior e inferior.
- El tejido mesófilo dentro de la hoja, que es rico en cloroplastos (también llamado clorénquima )
- La disposición de las venas (el tejido vascular)
Estos tres sistemas tisulares suelen formar una organización regular a escala celular. Las células especializadas que difieren notablemente de las células circundantes y que a menudo sintetizan productos especializados, como cristales, se denominan idioblastos.
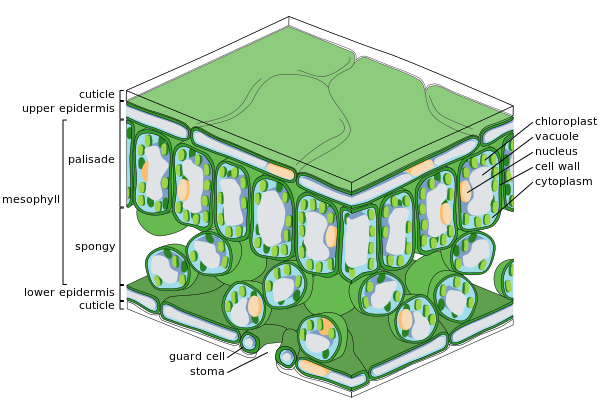
Principales tejidos de la hoja
 Sección transversal de una hoja
Sección transversal de una hoja Células epidérmicas
Células epidérmicas Células del mesófilo esponjoso
Células del mesófilo esponjoso
Epidermis
La epidermis es la capa externa de células que cubre la hoja. Está cubierto con una cutícula cerosa que es impermeable al agua líquida y al vapor de agua y forma el límite que separa las células internas de la planta del mundo externo. La cutícula es en algunos casos más delgada en la epidermis inferior que en la epidermis superior, y generalmente es más gruesa en las hojas de climas secos en comparación con las de climas húmedos. La epidermis cumple varias funciones: protección contra la pérdida de agua por transpiración, regulación del intercambio de gases y secreción de compuestos metabólicos. La mayoría de las hojas muestran una anatomía dorsoventral: las superficies superior (adaxial) e inferior (abaxial) tienen una construcción algo diferente y pueden cumplir funciones diferentes.
El tejido de la epidermis incluye varios tipos de células diferenciadas; células epidérmicas, células ciliadas epidérmicas (tricomas), células en el complejo estomático; celdas de guardia y celdas subsidiarias. Las células epidérmicas son las más numerosas, más grandes y menos especializadas y forman la mayor parte de la epidermis. Por lo general, son más alargadas en las hojas de las monocotiledóneas que en las de las dicotiledóneas.
Los cloroplastos generalmente están ausentes en las células epidérmicas, con la excepción de las células protectoras de los estomas. Los poros estomáticos perforan la epidermis y están rodeados a cada lado por células protectoras que contienen cloroplastos y de dos a cuatro células subsidiarias que carecen de cloroplastos, formando un grupo celular especializado conocido como complejo estomático. La apertura y cierre de la abertura estomática está controlada por el complejo estomático y regula el intercambio de gases y vapor de agua entre el aire exterior y el interior de la hoja. Por lo tanto, los estomas juegan un papel importante al permitir la fotosíntesis sin dejar que la hoja se seque. En una hoja típica, los estomas son más numerosos sobre la epidermis abaxial (inferior) que sobre la epidermis adaxial (superior) y son más numerosos en plantas de climas más fríos.
Mesófilo
La mayor parte del interior de la hoja entre las capas superior e inferior de la epidermis es un parénquima (tejido del suelo) o tejido clorénquima llamado mesófilo (en griego, "hoja media"). Este tejido de asimilación es la ubicación principal de la fotosíntesis en la planta. Los productos de la fotosíntesis se denominan "asimilados".
En los helechos y la mayoría de las plantas con flores, el mesófilo se divide en dos capas:
- Una capa superior en empalizada de células alargadas verticalmente, de una a dos células de espesor, directamente debajo de la epidermis adaxial, con espacios de aire intercelulares entre ellas. Sus células contienen muchos más cloroplastos que la capa esponjosa. Las células cilíndricas, con los cloroplastos cerca de las paredes de la célula, pueden aprovechar al máximo la luz. La ligera separación de las células proporciona la máxima absorción de dióxido de carbono. Las hojas de sol tienen una capa empalizada de varias capas, mientras que las hojas de sombra o las hojas más viejas más cercanas al suelo tienen una sola capa.
- Debajo de la capa empalizada se encuentra la capa esponjosa. Las células de la capa esponjosa están más ramificadas y no tan apretadas, de modo que hay grandes espacios de aire intercelulares entre ellas. Los poros o estomas de la epidermis se abren en cámaras subestomatales, que están conectadas a los espacios de aire intercelulares entre la célula mesófila esponjosa y empalizada, de modo que el oxígeno, el dióxido de carbono y el vapor de agua pueden difundirse dentro y fuera de la hoja y acceder a las células mesófilas. durante la respiración, la fotosíntesis y la transpiración.
Las hojas son normalmente verdes, debido a la clorofila en los cloroplastos en las células del mesófilo. Las plantas que carecen de clorofila no pueden realizar la fotosíntesis.
Tejido vascular
Las nervaduras son el tejido vascular de la hoja y se ubican en la capa esponjosa del mesófilo. El patrón de las venas se llama venación. En las angiospermas, la venación suele ser paralela en las monocotiledóneas y forma una red de interconexión en las plantas de hoja ancha. Alguna vez se pensó que eran ejemplos típicos de formación de patrones a través de la ramificación, pero en cambio pueden ejemplificar un patrón formado en un campo de tensor de tensión.
Una vena está formada por un haz vascular. En el núcleo de cada paquete hay grupos de dos tipos distintos de células conductoras:xilemaCélulas que llevan agua y minerales desde las raíces hasta la hoja.LíberCélulas que normalmente mueven la savia, con sacarosa disuelta (glucosa a sacarosa) producida por la fotosíntesis en la hoja, fuera de la hoja.
El xilema típicamente se encuentra en el lado adaxial del haz vascular y el floema típicamente se encuentra en el lado abaxial. Ambos están incrustados en un tejido de parénquima denso, llamado vaina, que generalmente incluye algo de tejido colénquima estructural.
Desarrollo de la hoja
De acuerdo con la teoría de brotes parciales de la hoja de Agnes Arber, las hojas son brotes parciales, que se derivan de los primordios de las hojas del ápice del brote. Al principio del desarrollo, se aplanan dorsiventralmente con las superficies dorsal y ventral. Las hojas compuestas están más cerca de los brotes que las hojas simples. Los estudios de desarrollo han demostrado que las hojas compuestas, como los brotes, pueden ramificarse en tres dimensiones. Sobre la base de la genética molecular, Eckardt y Baum (2010) concluyeron que "ahora se acepta generalmente que las hojas compuestas expresan propiedades tanto de hojas como de brotes".
Ecología
Biomecánica
Las plantas responden y se adaptan a los factores ambientales, como la luz y el estrés mecánico del viento. Las hojas necesitan sostener su propia masa y alinearse de tal manera que optimicen su exposición al sol, generalmente más o menos horizontalmente. Sin embargo, la alineación horizontal maximiza la exposición a las fuerzas de flexión y las fallas por estrés como el viento, la nieve, el granizo, la caída de escombros, los animales y la abrasión del follaje y las estructuras vegetales circundantes. En general, las hojas son relativamente endebles con respecto a otras estructuras de la planta, como tallos, ramas y raíces.
Tanto la lámina de la hoja como la estructura del pecíolo influyen en la respuesta de la hoja a fuerzas como el viento, lo que permite un grado de reposicionamiento para minimizar el arrastre y el daño, en lugar de la resistencia. El movimiento de la hoja como este también puede aumentar la turbulencia del aire cerca de la superficie de la hoja, lo que adelgaza la capa límite de aire inmediatamente adyacente a la superficie, lo que aumenta la capacidad de intercambio de gas y calor, así como la fotosíntesis. Las fuertes fuerzas del viento pueden dar como resultado una disminución del número de hojas y del área de superficie, lo que, si bien reduce la resistencia, implica una compensación por la reducción de la fotosíntesis. Por lo tanto, el diseño de la hoja puede implicar un compromiso entre la ganancia de carbono, la termorregulación y la pérdida de agua, por un lado, y el costo de soportar cargas tanto estáticas como dinámicas. En las plantas vasculares,
Muchas hojas dependen del soporte hidrostático dispuesto alrededor de un esqueleto de tejido vascular para su fuerza, que depende del mantenimiento del estado hídrico de la hoja. Tanto la mecánica como la arquitectura de la hoja reflejan la necesidad de transporte y soporte. Read y Stokes (2006) consideran dos modelos básicos, la forma "hidrostática" y la "hoja en I" (ver Fig. 1). Las hojas hidrostáticas, como las de Prostanthera lasianthos, son grandes y delgadas, y pueden implicar la necesidad de hojas múltiples en lugar de hojas grandes únicas debido a la cantidad de venas necesarias para sostener la periferia de las hojas grandes. Pero el tamaño grande de las hojas favorece la eficiencia en la fotosíntesis y la conservación del agua, lo que implica más compensaciones. Por otro lado, las hojas en I-beam como Banksia marginatainvolucrar estructuras especializadas para endurecerlos. Estas vigas en I se forman a partir de extensiones de la vaina del haz de esclerénquima que se encuentran con capas subepidérmicas rígidas. Esto cambia el equilibrio de la dependencia de la presión hidrostática al soporte estructural, una ventaja obvia donde el agua es relativamente escasa. Las hojas largas y angostas se doblan más fácilmente que las hojas ovadas de la misma área. Las monocotiledóneas suelen tener hojas lineales que maximizan el área de superficie y minimizan el sombreado propio. En estos, una alta proporción de venas principales longitudinales brindan soporte adicional.
Interacciones con otros organismos.
Aunque no son tan nutritivas como otros órganos como la fruta, las hojas proporcionan una fuente de alimento para muchos organismos. La hoja es una fuente vital de producción de energía para la planta, y las plantas han desarrollado protección contra los animales que consumen hojas, como los taninos, sustancias químicas que dificultan la digestión de las proteínas y tienen un sabor desagradable. Los animales que se especializan en comer hojas se conocen como folívoros.
Algunas especies tienen adaptaciones crípticas mediante las cuales usan hojas para evitar a los depredadores. Por ejemplo, las orugas de algunas polillas enrolladoras de hojas crean un pequeño hogar en la hoja doblándola sobre sí mismas. Algunas moscas de sierra enrollan de manera similar las hojas de sus plantas alimenticias en tubos. Las hembras de Attelabidae, los llamados gorgojos enrolladores de hojas, ponen sus huevos en las hojas que luego enrollan como medio de protección. Otros herbívoros y sus depredadores imitan la apariencia de la hoja. Los reptiles, como algunos camaleones, e insectos, como algunos saltamontes, también imitan los movimientos oscilantes de las hojas en el viento, moviéndose de un lado a otro o de un lado a otro mientras evaden una posible amenaza.
Pérdida estacional de hojas
Las hojas en las zonas templadas, boreales y estacionalmente secas pueden ser caducas estacionalmente (caerse o morir por las inclemencias de la estación). Este mecanismo para arrojar hojas se llama abscisión. Cuando se cae la hoja, deja una cicatriz de hoja en la ramita. En otoños fríos, a veces cambian de color y se vuelven amarillos, anaranjados brillantes o rojos, ya que varios pigmentos accesorios (carotenoides y xantofilas) se revelan cuando el árbol responde al frío y a la luz solar reducida reduciendo la producción de clorofila. Ahora se cree que los pigmentos de antocianina roja se producen en la hoja a medida que muere, posiblemente para enmascarar el tono amarillo que queda cuando se pierde la clorofila; las hojas amarillas parecen atraer a herbívoros como los pulgones.El enmascaramiento óptico de la clorofila por las antocianinas reduce el riesgo de daño fotooxidativo en las células de las hojas a medida que envejecen, lo que de otro modo podría reducir la eficiencia de la recuperación de nutrientes de las hojas otoñales envejecidas.
Adaptación evolutiva
En el curso de la evolución, las hojas se han adaptado a diferentes ambientes de las siguientes maneras:
- Las micro y nanoestructuras cerosas en la superficie reducen la humectación por la lluvia y la adhesión de la contaminación ( ver efecto Lotus ).
- Las hojas divididas y compuestas reducen la resistencia al viento y promueven el enfriamiento.
- Los pelos en la superficie de la hoja atrapan la humedad en climas secos y crean una capa límite que reduce la pérdida de agua.
- Las cutículas cerosas de las plantas reducen la pérdida de agua.
- El área de superficie grande proporciona un área grande para capturar la luz solar.
- En niveles nocivos de luz solar, las hojas especializadas, opacas o parcialmente enterradas, admiten la luz a través de una ventana de hoja translúcida para la fotosíntesis en las superficies internas de la hoja (por ejemplo, Fenestraria ).
- Anatomía de la hoja de Kranz en plantas que realizan la fijación de carbono C4
- Las hojas suculentas almacenan agua y ácidos orgánicos para su uso en la fotosíntesis CAM.
- Los aceites aromáticos, los venenos o las feromonas producidas por las glándulas de las hojas disuaden a los herbívoros (p. ej., eucaliptos).
- Las inclusiones de minerales cristalinos disuaden a los herbívoros (por ejemplo, fitolitos de sílice en pastos, rafidios en Araceae).
- Los pétalos atraen polinizadores.
- Las espinas protegen las plantas de los herbívoros (por ejemplo, cactus).
- Pelos punzantes para proteger contra la herbivoría, por ejemplo, en Urtica dioica y Dendrocnide moroides (Urticaceae).
- Las hojas especiales de las plantas carnívoras están adaptadas para atrapar alimentos, principalmente presas de invertebrados, aunque algunas especies también atrapan pequeños vertebrados (ver plantas carnívoras).
- Los bulbos almacenan alimentos y agua (p. ej., cebollas).
- Los zarcillos permiten que la planta trepe (p. ej., guisantes).
- Las brácteas y las pseudanthias (flores falsas) reemplazan las estructuras florales normales cuando las flores verdaderas están muy reducidas (p. ej., tártago, espatas en Araceae y capítulos florales en Asteraceae).
Terminología
Forma
Borde (margen)
| Imagen | Término | latín | Descripción |
|---|---|---|---|
| Entero | forma integra | Incluso; con un margen liso; sin dientes | |
| ciliado | ciliados | Flecos con pelos | |
| crenado | Crenata | de dientes ondulados; dentado con dientes redondeados | |
| Dentado | dentado | dentadoTal veztoscamente dentado, con dientes grandesodentado glandular, que tiene dientes que tienen glándulas | |
| denticulado | Denticulata | finamente dentado | |
| Doblemente aserrado | Duplicado-dentado | Cada diente con dientes más pequeños. | |
| Serrado | Serrata | Serrado; con dientes asimétricos apuntando hacia adelante | |
| serrulado | serrulata | finamente serrado | |
| Ondeado | Sinuoso | Con profundas hendiduras onduladas; toscamente almenado | |
| lóbulo | Lobatá | Con sangría, con las muescas que no llegan al centro. | |
| Ondular | Undulata | Con un borde ondulado, menos profundo que sinuoso | |
| espinoso oacre | espiguillas | Con puntas rígidas y afiladas como cardos |
Ápice (punta)
| Imagen | Término | latín | Descripción |
|---|---|---|---|
| Puntiagudo | _ | De punta larga, prolongada en una punta estrecha y ahusada de manera cóncava. | |
| Agudo | _ | Terminación en punta aguda, pero no prolongada. | |
| Con fines de cúspide | _ | Con una punta afilada, alargada y rígida; punta con una cúspide | |
| marginado | _ | Con sangría, con una muesca poco profunda en la punta | |
| mucronato | _ | Abruptamente inclinado con un pequeño punto corto | |
| mucronulado | _ | Mucronado, pero con una espina notablemente diminuta. | |
| obcordado | _ | Inversamente en forma de corazón | |
| Obtuso | _ | Redondeado o romo | |
| Truncar | _ | Terminando abruptamente con un extremo plano |
Base
PuntiagudoLlegando a un punto agudo, estrecho, prolongado.AgudoLlegando a un punto agudo, pero no prolongado.AuriculadoEn forma de oreja.En aspecto de corazónEn forma de corazón con la muesca hacia el tallo.PorcionesEn forma de cuña.AstadoCon forma de alabarda y con los lóbulos basales apuntando hacia afuera.OblicuoInclinación.reniformeCon forma de riñón pero más redonda y más ancha que larga.RedondeadoForma curva.FlechaCon forma de punta de flecha y con los lóbulos basales agudos apuntando hacia abajo.TruncarTerminando abruptamente con un extremo plano, que parece cortado.
Superficie
La superficie de la hoja también alberga una gran variedad de microorganismos; en este contexto se le conoce como la filosfera.lepidotaCubierto de finas escamas ásperas.
Lo peludo
Los "pelos" en las plantas se llaman propiamente tricomas. Las hojas pueden mostrar varios grados de vellosidad. El significado de varios de los siguientes términos puede superponerse.Aracnoides o aracnosisCon muchos pelos finos y enredados que le dan un aspecto de telaraña.BarbeladoCon pelos de púas finas (barbelas).BarbadoCon pelos largos y rígidos.ErizadoCon espinas rígidas como pelos.GrisáceoCanoso con densa pubescencia de color blanco grisáceo.ciliadoBordeado marginalmente con pelos cortos (cilios).CilioladoMinutamente ciliado.EscamosoCon bandadas de pelos suaves y lanudos, que tienden a desprenderse.glabrescentePérdida de cabello con la edad.GlabroNo hay pelos de ningún tipo presentes.GlandularCon una glándula en la punta del cabello.HirsutoCon pelos bastante ásperos o rígidos.híspidoCon pelos rígidos y erizados.hispíduloMinutamente híspido.CanosoCon una fina pubescencia cerrada de color blanco grisáceo.Lanato o lanosaCon pelos lanudos.PiloseCon pelos suaves y claramente separados.Puberulento o pubéruloCon pelos finos y diminutos.PubescenteCon pelos suaves, cortos y erectos.Escabroso o escabrosoÁspero al tacto.SedosoAspecto sedoso a través de pelos finos, rectos y adpresos (juntos y planos).SedosoCon pubescencia adprimida, suave y recta.estrellado o esteliformeCon pelos en forma de estrella.StrigoCon pelos adpresos, afilados, rectos y rígidos.tomentosoDensamente pubescente con pelos lanudos blancos suaves y enmarañados.Cano-tomentosaEntre canescente y tomentosa.Fieltro-tomentosaLanoso y enmarañado con pelos rizados.tomentulosaMinuciosamente o sólo ligeramente tomentoso.VellosoCon pelos largos y suaves, generalmente curvos.LanosoCon pelos largos, suaves y tortuosos o enmarañados.
Sincronización
histeránticoDesarrollando después de las flores.sinantosoDesarrollándose al mismo tiempo que las flores.
Venación
Clasificación
Se han descrito varios sistemas de clasificación diferentes de los patrones de las nervaduras de las hojas (venación o nervadura), comenzando con Ettingshausen (1861), junto con muchos términos descriptivos diferentes, y la terminología se ha descrito como "formidable". Uno de los más comunes entre estos es el sistema Hickey, desarrollado originalmente para "dicotiledóneas" y que usa varios términos de Ettingshausen derivados del griego (1973-1979): ( ver también : Simpson Figura 9.12, p. 468)
Sistema de chupetón
1. Pinnadas (nervadas como plumas, reticuladas, en red pinnadas, acanaladas, acanaladas o nervadas)Las venas surgen pinnadas (como plumas) de una sola vena primaria (vena media) y se subdividen en venillas secundarias, conocidas como venas de orden superior. Estos, a su vez, forman una red complicada. Este tipo de venación es típico de (pero de ninguna manera limitado a) "dicotiledóneas" (angiospermas no monocotiledóneas). Por ejemplo, Ostrya.Hay tres subtipos de venación pinnada:Craspedodromous (griego: kraspedon - borde, dromos - correr)Las venas principales llegan hasta el margen de la hoja.camptódromoLas venas principales se extienden cerca del margen, pero se doblan antes de cruzarse con el margen.HidrómoTodas las venas secundarias están ausentes, son rudimentarias o están ocultasEstos, a su vez, tienen una serie de subtipos adicionales, como eucamptódromos, donde las venas secundarias se curvan cerca del margen sin unirse a las venas secundarias adyacentes.Pinada

craspedódromo

camptódromo

Hidrómo2. Paralelodromo (paralelo-nervado, paralelo-nervado, paralelo-nervado, penniparalelo, estriado)Dos o más nervios primarios que se originan uno al lado del otro en la base de la hoja y corren paralelos entre sí hasta el ápice y luego convergen allí. Las venas comisurales (venas pequeñas) conectan las principales venas paralelas. Típico de la mayoría de las monocotiledóneas, como las gramíneas.También se utilizan los términos adicionales marginal (las venas primarias alcanzan el margen) y reticulado (con venas netas).Paralelodromo

3. Campylodromous ( campylos - curva)Varias venas o ramas primarias que se originan en o cerca de un solo punto y corren en arcos recurvados, luego convergen en el vértice. Por ejemplo Maianthemum.campilódromo

4. AcródromoDos o más nervios primarios o secundarios bien desarrollados en arcos convergentes hacia el ápice, sin recurvatura basal como en Campilódromo. Pueden ser basales o suprabasales según el origen, y perfectos o imperfectos según lleguen a los 2/3 del camino al ápice. Por ejemplo, Miconia (tipo basal), Endlicheria (tipo suprabasal).acródromo

Basal imperfecto

Suprabasal imperfecto

Basal perfecto

Suprabasal perfecto5. ActinódromoTres o más venas primarias que divergen radialmente desde un solo punto. Por ejemplo, Arcangelisia (tipo basal), Givotia (tipo suprabasal).actinódromo

Imperfecto marginal

Imperfecto reticulado6. PalinactódromoNervios primarios con uno o más puntos de ramificación dicotómica secundaria más allá de la divergencia primaria, ya sea más cercanos o más distantes. Por ejemplo, Platano.palinactódromo

Los tipos 4 a 6 pueden subclasificarse de manera similar como basales (primarias unidas en la base de la hoja) o suprabasales (divergiendo sobre la base de la hoja), y perfectos o imperfectos, pero también flabelados.
Casi al mismo tiempo, Melville (1976) describió un sistema aplicable a todas las angiospermas y utilizando terminología latina e inglesa. Melville también tenía seis divisiones, según el orden en que se desarrollan las venas.Arbuscular (arbuscularis)Ramificación repetida por dicotomía regular para dar lugar a una estructura similar a un arbusto tridimensional que consiste en un segmento lineal (2 subclases)Fanbellate (abanico)Nervios primarios rectos o solo ligeramente curvados, divergiendo de la base en forma de abanico (4 subclases)Palmato (palmatus)Venas primarias curvas (3 subclases)Pinnada (pinnatus)Vena primaria única, la nervadura central, a lo largo de la cual se disponen venas secundarias rectas o arqueadas a intervalos más o menos regulares (6 subclases)colimar (collimatus)Numerosas venas primarias longitudinalmente paralelas que surgen de un meristemo transversal (5 subclases)Conglutinar (conglutinatus)Derivado de folletos pinnados fusionados (3 subclases)
Posteriormente, se incorporó una forma modificada del sistema Hickey a la clasificación Smithsonian (1999), que proponía siete tipos principales de venación, según la arquitectura de las venas primarias, y agregó Flabellate como un tipo principal adicional. Luego se hizo una clasificación adicional sobre la base de las venas secundarias, con 12 tipos más, como;broquidódromoForma cerrada en la que las secundarias se unen en una serie de arcos prominentes, como en Hildegardia.craspedódromoForma abierta con secundarias que terminan en el margen, en hojas dentadas, como en Celtis.EucamptódromoForma intermedia con secundarias hacia arriba que disminuyen gradualmente apicalmente pero dentro del margen, y conectadas por venas terciarias intermedias en lugar de bucles entre secundarias, como en Cornus.cladodromoSecundarias que se ramifican libremente hacia el margen, como en Rhus.
términos que se habían utilizado como subtipos en el sistema Hickey original.Patrones de venación secundaria

broquidódromo

craspedódromo

Eucamptódromo

cladodromo

Broquidódromo
Hildegardia migeodii

Craspedódromo
Celtis occidentalis

Cornus officinalis eucamptódromo

Cladódromo
Rhus ovata
Otras descripciones incluyeron el orden superior o las venas menores y los patrones de las areolas ( ver Grupo de trabajo de arquitectura de hojas, Figuras 28–29).faneladoVarias a muchas venas basales finas iguales que divergen radialmente en ángulos bajos y se ramifican apicalmente. Por ejemplo, Paranomus.fanelado

Los análisis de los patrones de vetas a menudo se toman en consideración del orden de las vetas, el tipo de veta principal, el tipo de veta secundaria (venas principales) y la densidad de las venas menores. Varios autores han adoptado versiones simplificadas de estos esquemas. En su forma más simple, los tipos de vetas primarias se pueden considerar en tres o cuatro grupos según las divisiones de plantas que se consideren;
- pinada
- palmeado
- paralelo
donde palmeado se refiere a múltiples venas primarias que irradian desde el pecíolo, en oposición a la ramificación de la vena principal central en forma pinnada, y abarca los tipos 4 y 5 de Hickey, que se conservan como subtipos; por ejemplo, palmeado-acródromo ( consulte la Guía de hojas del Servicio de Parques Nacionales).Palmate, Palmate-netted, palmate-veined, fan-vetedVarias venas principales de aproximadamente el mismo tamaño divergen desde un punto común cerca de la base de la hoja donde se une el pecíolo y se irradian hacia el borde de la hoja. Las hojas palmeadas a menudo son lobuladas o divididas con lóbulos que irradian desde el punto común. Pueden variar en el número de venas primarias (3 o más), pero siempre irradian desde un punto común. por ejemplo, la mayoría de Acer (arces).Palmeado

Otros sistemas
Alternativamente, Simpson usa:sin nerviosNervio central central sin venas laterales (micrófilo), visto en las traqueofitas que no tienen semillas, como las colas de caballodicotómicoVenas que se ramifican sucesivamente en venas de igual tamaño desde un punto común, formando una unión en Y, abriéndose en abanico. Entre las plantas leñosas templadas, Ginkgo biloba es la única especie que exhibe venación dicotómica. También algunos pteridofitos (helechos).ParaleloLas venas primarias y secundarias son aproximadamente paralelas entre sí, a lo largo de la hoja, a menudo conectadas por enlaces perpendiculares cortos, en lugar de formar redes. En algunas especies, las venas paralelas se unen en la base y el ápice, como los árboles de hoja perenne y las gramíneas. Característica de las monocotiledóneas, pero las excepciones incluyen Arisaema y, como se muestra a continuación, debajo de la red.Enredado (reticulado, pinnado)Una vena central prominente con venas secundarias que se ramifican a ambos lados. El nombre deriva de las vetas últimas que forman una red interconectada como patrón o red. (La nervadura primaria y secundaria puede denominarse pinnada, mientras que las nervaduras más finas en forma de red se denominan enredadas o reticuladas); la mayoría de las angiospermas no monocotiledóneas, con excepciones como Calophyllum. Algunas monocotiledóneas tienen venación reticulada, incluidas Colocasia, Dioscorea y Smilax.

Equisetum :Hojas micrófilas reducidas (L) que surgen en verticilos desde el nudo

Ginkgo biloba :venación dicotómica
Sin embargo, estos sistemas simplificados permiten una mayor división en múltiples subtipos. Simpson, (y otros) divide paralelo y neto (y algunos utilizan sólo estos dos términos para las angiospermas) sobre la base del número de venas primarias (costa) de la siguiente manera;ParaleloPenni-paralelo (pinnado, pinnado paralelo, unicostado paralelo)Una sola nervadura central central prominente, las venas secundarias de esta surgen perpendicularmente y corren paralelas entre sí hacia el margen o la punta, pero no se unen (anastomosan). El término unicostado se refiere a la prominencia de la única nervadura central (costa) que se extiende a lo largo de la hoja desde la base hasta el ápice. por ejemplo, Zingiberales, como plátanos, etc.Palmate-paralelo (multicostate paralelo)Varias venas primarias igualmente prominentes que surgen de un solo punto en la base y corren paralelas hacia la punta o el margen. El término multicostado se refiere a tener más de una vena principal prominente. por ejemplo, palmeras "abanico" (palmadas) (Arecaceae)Convergente paralelo multiestadoLas venas medias convergen en el ápice, por ejemplo, Bambusa arundinacea = B. bambos (Aracaceae), EichorniaParalelo multicostado divergenteLas nervaduras medias divergen más o menos paralelas hacia el margen, por ejemplo, Borassus (Poaceae), palmitosNetted (reticulado)Pinnado (nervado, enredado, reticulado unicostato)Una sola nervadura central prominente que se extiende desde la base hasta el ápice, las venas secundarias se originan a ambos lados a lo largo de la nervadura principal principal y corren hacia el margen o ápice (punta), con una red de venillas más pequeñas que forman un retículo (malla o red). por ejemplo, Mangifera, Ficus religiosa, Psidium guajava, Hibiscus rosa-sinensis, Salix albaPalmately (multicostate reticulate)Más de una vena primaria que surge de un solo punto, que va desde la base hasta el ápice. por ejemplo, Liquidambar styraciflua Esto puede subdividirse aún más;Convergente de múltiples estadosLas venas principales divergen desde el origen en la base y luego convergen hacia la punta. por ejemplo, Zizyphus, Smilax, Cinnamomumdivergente multicostatalTodas las venas principales divergen hacia la punta. por ejemplo, Gossypium, Cucurbita, Carica papaya, Ricinus communisTernately (ternate-neted)Tres venas primarias, como arriba, p. ej. ( ver ) Ceanothus leucodermis, C. tomentosus, Encelia farinosaPatrones de venación de Simpson

Maranta leuconeura var.
erythroneura (Zingiberales):Penni-paralelo

Coccothrinax argentea (Arecaceae):Palmate-paralelo

Bambusa bambos :Multicostate paralelo convergente

Borassus sp.:Paralelo divergente multicostado
Salix alba :Redada pinnadamente

Liquidambar styraciflua :palmeada

Ziziphus jujuba :convergente palmeada multicostada

Gossypium tomentosum :divergente palmeada multicostada
Estos sistemas complejos no se utilizan mucho en las descripciones morfológicas de taxones, pero tienen utilidad en la identificación de plantas, aunque se critican por estar excesivamente cargados de jerga.
Un sistema más antiguo, incluso más simple, utilizado en alguna flora utiliza solo dos categorías, abierta y cerrada.
- Abierto: Las venas de orden superior tienen terminaciones libres entre las células y son más características de las angiospermas no monocotiledóneas. Es más probable que se asocien con formas de hojas dentadas, lobuladas o compuestas. Pueden subdividirse como;
- Hojas pinnadas (nervadas como plumas), con una nervadura central principal o nervadura (nervadura central), de la que surge el resto del sistema de nervadura
- Palmate, en el que tres o más costillas principales se elevan juntas en la base de la hoja y divergen hacia arriba.
- Dicotómico, como en los helechos, donde las venas se bifurcan repetidamente
- Cerrado: Las venas de orden superior están conectadas en bucles sin terminar libremente entre las células. Estos tienden a estar en hojas con contornos suaves y son característicos de las monocotiledóneas.
- Se pueden subdividir en si las venas corren paralelas, como en los pastos, o tienen otros patrones.
Otros términos descriptivos
También hay muchos otros términos descriptivos, a menudo con un uso muy especializado y limitados a grupos taxonómicos específicos. La visibilidad de las venas depende de una serie de características. Estos incluyen el ancho de las venas, su prominencia en relación con la superficie de la lámina y el grado de opacidad de la superficie, que puede ocultar venas más finas. En este sentido, las venas se denominan oscuras y el orden de las venas que están oscurecidas y si la superficie superior, la inferior o ambas, se especifica con más detalle.
Los términos que describen la prominencia de la vena incluyen bullado, canalizado, plano, acanalado, impreso, prominente y hundido ( Fig. 6.1 Hawthorne & Lawrence 2013). Las venas pueden mostrar diferentes tipos de prominencia en diferentes áreas de la hoja. Por ejemplo, Pimenta racemosa tiene una nervadura central acanalada en la superficie superior, pero esto es prominente en la superficie inferior.
Describiendo la prominencia de la vena:BullateSuperficie de la hoja levantada en una serie de cúpulas entre las nervaduras en la cara superior, y por lo tanto también con marcadas depresiones. por ejemplo, Rytigynia pauciflora, Vitis viniferaCanalizado (canalicululado)Venas hundidas debajo de la superficie, dando como resultado un canal redondeado. A veces se confunde con "canalizado" porque los canales pueden funcionar como canaletas para que la lluvia se escurra y permita el secado, como en muchas Melastomataceae. por ejemplo ( ver ) Pimenta racemosa (Myrtaceae), Clidemia hirta (Melastomataceae).CanalonesNervios parcialmente prominentes, la cresta sobre la superficie de la lámina de la hoja, pero con canales a lo largo de cada lado, como canalesImpresionadoVena que forma una línea elevada o una cresta que se encuentra debajo del plano de la superficie que la sostiene, como si estuviera presionada contra ella, y que a menudo está expuesta en la superficie inferior. El tejido cerca de las venas a menudo parece arrugarse, dándoles una apariencia hundida o en relieve.OscuroVenas no visibles o nada claras; si no se especifica, entonces no es visible a simple vista. por ejemplo, Berberis gagnepainii. En este Berberis, las venas solo son oscuras en la superficie inferior.ProminenteVena elevada por encima de la superficie circundante para que se sienta fácilmente cuando se acaricia con el dedo. por ejemplo ( ver ) Pimenta racemosa, Spathiphyllum cannifoliumEmpotradoLa vena está hundida debajo de la superficie, más prominente que los tejidos circundantes pero más hundida en el canal que con las venas impresas. por ejemplo, Viburnum plicatum.Tipos de prominencia de la vena
Vitis vinífera
Bullate
Clidemia hirta
canalizada

cornejo
impresionado

Berberis gagnepainii
Oscuro (bajo la superficie)

Spathiphyllum cannifolium
Destacado

Viburnum plicatum
Empotrado
Describiendo otras características:Plinervy (plinerved)Más de una vena principal (nervio) en la base. Nervios secundarios laterales que se ramifican desde un punto por encima de la base de la hoja. Por lo general, se expresa como un sufijo, como en una hoja de tres o tres hojas. En una hoja de 3 plinerved (triplinerved), tres venas principales se ramifican por encima de la base de la lámina (dos venas secundarias y la vena principal) y corren esencialmente paralelas posteriormente, como en Ceanothus y en Celtis. De manera similar, una hoja quintuplinerve (cinco venas) tiene cuatro venas secundarias y una vena principal. Un patrón con 3-7 venas es especialmente llamativo en Melastomataceae. El término también se ha utilizado en Vaccinieae. El término se ha utilizado como sinónimo de acródromo, palmato-acródromo o suprabasal, y se cree que su definición es demasiado amplia.escalariformeVenas dispuestas como los peldaños de una escalera, particularmente venas de orden superiorsubmarginalVenas corriendo cerca del margen de la hojatrinervado2 nervios basales principales además de la nervadura central
Diagramas de patrones de venación.
| Imagen | Término | Descripción |
|---|---|---|
 | Arqueado | Arco secundario hacia el ápice. |
 | dicotómico | Venas partidas en dos |
 | Longitudinal | Todas las venas alineadas principalmente con la vena media |
 | Paralelo | Todas las venas son paralelas y no se cruzan. |
 | Pinada | Venas secundarias que nacen de la nervadura central |
 | Reticular | Todas las venas se ramifican repetidamente, venas netas |
 | Girar | Venas que salen del centro de la hoja y se irradian hacia los bordes. |
 | Transverso | Venas terciarias que corren perpendiculares al eje de la vena principal, conectando venas secundarias |
Tamaño
Los términos megafila, macrófila, mesófila, notófila, micrófila, nanofila y leptofila se utilizan para describir el tamaño de las hojas (en orden descendente), en una clasificación ideada en 1934 por Christen C. Raunkiær y modificada desde entonces por otros.
Contenido relacionado
Epidermis (botánica)