
Equinodermo
Un equinodermo () es cualquier miembro del phylum Echinodermata (). Los adultos son reconocibles por su simetría radial (generalmente de cinco puntos) e incluyen estrellas de mar, estrellas frágiles, erizos de mar, dólares de arena y pepinos de mar, así como lirios de mar o "lirios de piedra". Los equinodermos adultos se encuentran en el lecho marino en todas las profundidades del océano, desde la zona intermareal hasta la zona abisal. El filo contiene unas 7.000 especies vivas, lo que lo convierte en el segundo grupo más grande de deuteróstomos, después de los cordados. Los equinodermos son el filo completamente marino más grande. Los primeros equinodermos definitivos aparecieron cerca del comienzo del Cámbrico.
Los equinodermos son importantes tanto ecológica como geológicamente. Ecológicamente, hay pocas otras agrupaciones tan abundantes en el desierto biótico de las profundidades marinas, así como en los océanos menos profundos. La mayoría de los equinodermos pueden reproducirse asexualmente y regenerar tejidos, órganos y extremidades; en algunos casos, pueden sufrir una regeneración completa a partir de una sola extremidad. Geológicamente, el valor de los equinodermos está en sus esqueletos osificados, que son los principales contribuyentes a muchas formaciones de piedra caliza y pueden proporcionar pistas valiosas sobre el entorno geológico. Fueron las especies más utilizadas en la investigación regenerativa en los siglos XIX y XX. Además, algunos científicos sostienen que la radiación de los equinodermos fue responsable de la Revolución Marina Mesozoica.
Taxonomía y evolución
El nombre equinodermo proviene del griego antiguo ἐχῖνος (ekhînos) 'erizo', y δέρμα (dérma) 'piel'. Los equinodermos son bilaterales, lo que significa que sus ancestros tenían simetría especular. Entre los bilaterales, pertenecen a la división deuterostoma, lo que significa que el blastoporo, la primera abertura que se forma durante el desarrollo del embrión, se convierte en el ano en lugar de la boca. Las características de los equinodermos adultos son la posesión de un sistema vascular acuático con pies ambulacrales externos y un endoesqueleto calcáreo formado por huesecillos conectados por una malla de fibras de colágeno.
Filogenia
Históricamente, los taxónomos creían que Ophiuroidea era hermana de Asteroidea, o que eran hermanas de (Holothuroidea + Echinoidea). Sin embargo, un análisis de 2014 de 219 genes de todas las clases de equinodermos revisó el árbol filogenético. Un análisis independiente en 2015 de transcriptomas de ARN de 23 especies en todas las clases de equinodermos dio el mismo árbol.
- Fylogeny externa
El contexto de los equinodermos dentro de Bilateria es:
| Bilateria |
| ||||||||||||||||||||||||||||||
.jpg)
- Filogenia interna
| Echino‐ |
| ||||||||||||||||||||||||
| dermata |

Diversidad
Existen unas 7000 especies de equinodermos, así como unas 13 000 especies extintas. Todos los equinodermos son marinos, pero se encuentran en hábitats que van desde áreas intermareales poco profundas hasta profundidades abisales. Tradicionalmente se reconocen dos subdivisiones principales: los Eleutherozoa móviles más familiares, que abarcan Asteroidea (estrellas de mar, con unas 1745 especies), Ophiuroidea (estrellas frágiles, con unas 2300 especies), Echinoidea (erizos de mar y dólares de arena, con unas 900 especies) y Holothuroidea (pepinos de mar, con unas 1.430 especies); y los Pelmatozoa, algunos de los cuales son sésiles mientras que otros son móviles. Estos consisten en Crinoidea (estrellas de plumas y lirios de mar, con alrededor de 580 especies) y los blastoides y Paracrinoideos extintos.
- La subfila de los echinodermos

Una estrella frágil, Ophionereis reticulata

Un pepino de mar de Malasia

Starfish de colores variados

Un erizo de mar, Strongylocentrotus purpuratus

Crinoide en un arrecife de coral




Historia de los fósiles
El fósil de equinodermo candidato más antiguo es Arkarua del Precámbrico de Australia. Estos fósiles tienen forma de disco, con crestas radiales en el borde y una depresión central de cinco puntas marcada con líneas radiales. Sin embargo, los fósiles no tienen estereoma o estructura interna que indique un sistema vascular de agua, por lo que no pueden identificarse de manera concluyente.
Los primeros equinodermos universalmente aceptados aparecen en el período Cámbrico Inferior; los asterozoos aparecieron en el Ordovícico, mientras que los crinoideos fueron un grupo dominante en el Paleozoico. Los equinodermos dejaron un extenso registro fósil. Se plantea la hipótesis de que el antepasado de todos los equinodermos era un animal simple, móvil, bilateralmente simétrico, con boca, intestino y ano. Este organismo ancestral adoptó un modo de vida adjunto con alimentación en suspensión y desarrolló simetría radial. Aun así, las larvas de todos los equinodermos son simétricas bilateralmente y todas desarrollan simetría radial en la metamorfosis. Al igual que su antepasado, las estrellas de mar y los crinoideos todavía se adhieren al lecho marino mientras cambian a su forma adulta.
Los primeros equinodermos no tenían movilidad, pero evolucionaron hasta convertirse en animales capaces de moverse libremente. Estos pronto desarrollaron placas endoesqueléticas con estructura estéreo y surcos ciliares externos para alimentarse. Los equinodermos del Paleozoico eran globulares, adheridos al sustrato y estaban orientados con sus superficies orales hacia arriba. Estos primeros equinodermos tenían surcos ambulacrales que se extendían por el costado del cuerpo, bordeados a ambos lados por braquiolos, como las pínnulas de un crinoideo moderno. Eventualmente, a excepción de los crinoideos, todas las clases de equinodermos invirtieron su orientación para volverse boca abajo. Antes de que esto sucediera, los podios probablemente tenían una función de alimentación, como lo hacen hoy en día en los crinoideos. La función locomotora de los podios vino más tarde, cuando la reorientación de la boca puso los podios en contacto con el sustrato por primera vez.
- Más información: Dibrachicystis
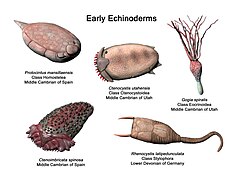
Equinodermos tempranos Ctenoimbricata, Ctenocystis, Gogia, Protocintus y Rhenocystis

El citoide ordoviciano Echinosphaerites del noreste de Estonia

Coronas crinoides fossil

Cáliz de Hyperoblastus, un blastoid del Devonian de Wisconsin




Anatomía y fisiología
Los equinodermos evolucionaron de animales con simetría bilateral. Aunque los equinodermos adultos poseen simetría pentaradial, sus larvas son organismos ciliados que nadan libremente con simetría bilateral. Más tarde, durante la metamorfosis, el lado izquierdo del cuerpo crece a expensas del lado derecho, que finalmente se absorbe. Luego, el lado izquierdo crece en forma de simetría pentarradial, en la que el cuerpo se organiza en cinco partes alrededor de un eje central. Dentro de los Asterozoa, puede haber algunas excepciones a la regla. La mayoría de las estrellas de mar del género Leptasterias tienen seis brazos, aunque pueden aparecer ejemplares de cinco brazos. Los Brisingida también contienen algunas especies de seis brazos. Entre las estrellas frágiles, existen especies de seis brazos como Ophiothela danae, Ophiactis savignyi y Ophionotus hexactis, y Ophiacantha vivipara a menudo tiene más de seis.
Los equinodermos tienen simetría radial secundaria en partes de su cuerpo en alguna etapa de la vida, muy probablemente una adaptación a una existencia sésil o de movimiento lento. Muchos crinoideos y algunas estrellas de mar son simétricos en múltiplos de los cinco básicos; las estrellas de mar como Labidiaster annulatus poseen hasta cincuenta brazos, mientras que el lirio de mar Comaster schlegelii tiene doscientos.
Piel y esqueleto
Los equinodermos tienen un esqueleto mesodérmico en la dermis, compuesto por placas a base de calcita conocidas como osículos. Si son sólidos, formarían un esqueleto pesado, por lo que tienen una estructura porosa similar a una esponja conocida como stereom. Los huesecillos pueden fusionarse, como en la prueba de los erizos de mar, o pueden articularse para formar articulaciones flexibles como en los brazos de las estrellas de mar, las estrellas frágiles y los crinoideos. Los huesecillos pueden tener proyecciones externas en forma de espinas, gránulos o verrugas y están sostenidos por una epidermis resistente. Los elementos esqueléticos a veces se despliegan de formas especializadas, como el órgano de masticación llamado 'linterna de Aristóteles'. en los erizos de mar, los tallos de apoyo de los crinoideos y el "anillo de cal" estructural; de pepinos de mar.
Aunque los huesecillos individuales son robustos y se fosilizan fácilmente, los esqueletos completos de estrellas de mar, estrellas frágiles y crinoideos son raros en el registro fósil. Por otro lado, los erizos de mar suelen estar bien conservados en lechos de yeso o piedra caliza. Durante la fosilización, las cavidades del estereoma se rellenan con calcita que se continúa con la roca circundante. Al fracturar dicha roca, los paleontólogos pueden observar patrones distintivos de clivaje y, a veces, incluso la intrincada estructura interna y externa de la prueba.
La epidermis contiene células pigmentarias que proporcionan los colores a menudo vívidos de los equinodermos, que incluyen rojo intenso, rayas blancas y negras y púrpura intenso. Estas células pueden ser sensibles a la luz, lo que hace que muchos equinodermos cambien de apariencia por completo cuando cae la noche. La reacción puede ocurrir rápidamente: el erizo de mar Centrostephanus longispinus cambia de color en tan solo cincuenta minutos cuando se expone a la luz.
Una característica de la mayoría de los equinodermos es un tipo especial de tejido conocido como tejido conectivo catch. Este material a base de colágeno puede cambiar sus propiedades mecánicas bajo el control nervioso más que por medios musculares. Este tejido permite que una estrella de mar pase de moverse con flexibilidad por el fondo marino a volverse rígida mientras abre un molusco bivalvo o evita que la extraigan de una grieta. De manera similar, los erizos de mar pueden bloquear sus espinas normalmente móviles en posición vertical como mecanismo defensivo cuando son atacados.
El sistema vascular del agua

Los equinodermos poseen un sistema vascular de agua único, una red de canales llenos de líquido derivados del celoma (cavidad corporal) que funcionan en el intercambio de gases, la alimentación, la recepción sensorial y la locomoción. Este sistema varía entre las diferentes clases de equinodermos, pero generalmente se abre al exterior a través de una madreporita similar a un tamiz en la superficie aboral (superior) del animal. La madreporita está unida a un conducto delgado, el canal de piedra, que se extiende hasta un canal anular que rodea la boca o el esófago. El canal anular se ramifica en un conjunto de canales radiales, que en los asteroides se extienden a lo largo de los brazos, y en los equinoides se unen a la prueba en las áreas ambulacrales. Los canales laterales cortos se ramifican de los canales radiales, cada uno de los cuales termina en una ampolla. Parte de la ampolla puede sobresalir a través de un poro (o un par de poros en los erizos de mar) hacia el exterior, formando un podio o pie de tubo. El sistema vascular de agua ayuda con la distribución de nutrientes por todo el cuerpo del animal; es más visible en los pies ambulacrales que pueden extenderse o contraerse por la redistribución de líquido entre el pie y la ampolla interna.
La organización del sistema vascular del agua es algo diferente en los ofiuroides, donde la madreporita puede estar en la superficie oral y los podios carecen de ventosas. En los holoturoideos, el sistema se reduce, a menudo con pocos pies tubulares además de los tentáculos de alimentación especializados, y la madreporita se abre hacia el celoma. Algunos holoturoides como Apodida carecen de pies ambulacrales y canales a lo largo del cuerpo; otros tienen canales longitudinales. La disposición en los crinoideos es similar a la de los asteroides, pero los pies tubulares carecen de ventosas y se utilizan en un movimiento de ida y vuelta para pasar las partículas de comida capturadas por los brazos hacia la boca central. En los asteroides, se emplea el mismo movimiento para mover al animal por el suelo.
Otros órganos
Los equinodermos poseen un sistema digestivo simple que varía según la dieta del animal. Las estrellas de mar son en su mayoría carnívoras y tienen boca, esófago, estómago, intestino y recto en dos partes, con el ano ubicado en el centro de la superficie corporal aboral. Con algunas excepciones, los miembros de la orden Paxillosida no poseen ano. En muchas especies de estrellas de mar, el gran estómago cardíaco puede evertirse para digerir alimentos fuera del cuerpo. Algunas otras especies pueden ingerir alimentos enteros como moluscos. Las estrellas frágiles, que tienen dietas variadas, tienen un intestino ciego sin intestino ni ano; expulsan los restos de comida por la boca. Los erizos de mar son herbívoros y utilizan sus piezas bucales especializadas para pastar, desgarrar y masticar su comida, principalmente algas. Tienen un esófago, un estómago grande y un recto con el ano en el vértice de la prueba. Los pepinos de mar son en su mayoría detritívoros, clasificando el sedimento con patas de tubo modificadas alrededor de la boca, los tentáculos bucales. La arena y el barro acompañan su alimentación a través de su tripa simple, que tiene un intestino largo y enrollado y una cloaca grande. Los crinoideos se alimentan de suspensión, atrapando pasivamente el plancton que se desplaza hacia sus brazos extendidos. Los bolos de comida atrapada en la mucosidad se pasan a la boca, que está unida al ano por un asa que consta de un esófago corto y un intestino más largo.
Las cavidades celómicas de los equinodermos son complejas. Aparte del sistema vascular de agua, los equinodermos tienen un celoma hemal, un celoma perivisceral, un celoma gonadal y, a menudo, también un celoma perihemal. Durante el desarrollo, el celoma del equinodermo se divide en metacoel, mesocoel y protocoel (también llamados somatocoel, hydrocoel y axocoel, respectivamente). El sistema vascular de agua, el sistema hemal y el sistema perihemal forman el sistema celómico tubular. Los equinodermos son inusuales porque tienen un sistema circulatorio celómico (el sistema vascular del agua) y un sistema circulatorio hemal, ya que la mayoría de los grupos de animales tienen solo uno de los dos.
Los sistemas hemal y perihemal se derivan del celoma original, formando un sistema circulatorio abierto y reducido. Por lo general, consta de un anillo central y cinco vasos radiales. No existe un verdadero corazón, y la sangre a menudo carece de cualquier pigmento respiratorio. El intercambio gaseoso se produce a través de branquias dérmicas o pápulas en estrellas de mar, bursas genitales en estrellas frágiles, branquias peristominales en erizos de mar y árboles cloacales en pepinos de mar. El intercambio de gases también tiene lugar a través de los pies tubulares. Los equinodermos carecen de órganos excretores (eliminación de desechos) especializados y, por lo tanto, los desechos nitrogenados, principalmente en forma de amoníaco, se difunden a través de las superficies respiratorias.
El fluido celómico contiene los celomocitos o células inmunitarias. Hay varios tipos de células inmunitarias, que varían entre clases y especies. Todas las clases poseen un tipo de amebocitos fagocíticos, que engullen partículas invasoras y células infectadas, se agregan o coagulan y pueden participar en la citotoxicidad. Estas células suelen ser grandes y granulares, y se cree que son una línea principal de defensa contra patógenos potenciales. Dependiendo de la clase, los equinodermos pueden tener células esféricas (para citotoxicidad, inflamación y actividad antibacteriana), células vibrables (para el movimiento y la coagulación del fluido celómico) y células cristalinas (que pueden servir para la osmorregulación en pepinos de mar). Los celomocitos secretan péptidos antimicrobianos contra bacterias, y cuentan con un conjunto de lectinas y proteínas del complemento como parte de un sistema inmunitario innato que aún está en proceso de caracterización.
Los equinodermos tienen un sistema nervioso radial simple que consta de una red nerviosa modificada de neuronas interconectadas sin cerebro central, aunque algunos poseen ganglios. Los nervios se irradian desde los anillos centrales alrededor de la boca hacia cada brazo oa lo largo de la pared del cuerpo; las ramas de estos nervios coordinan los movimientos del organismo y la sincronización de los pies ambulacrales. Las estrellas de mar tienen células sensoriales en el epitelio y tienen manchas oculares simples y pies de tubo similares a tentáculos sensibles al tacto en las puntas de sus brazos. Los erizos de mar no tienen órganos sensoriales particulares, pero tienen estatocistos que ayudan en la orientación gravitacional, y tienen células sensoriales en su epidermis, particularmente en los pies ambulacrales, las espinas dorsales y los pedicelarios. Las estrellas frágiles, los crinoideos y los pepinos de mar en general no tienen órganos sensoriales, pero algunos pepinos de mar excavadores del orden Apodida tienen un solo estatocisto junto a cada nervio radial, y algunos tienen una mancha ocular en la base de cada tentáculo.
Las gónadas ocupan, al menos periódicamente, gran parte de las cavidades corporales de los erizos de mar y los pepinos de mar, mientras que los crinoideos, las estrellas frágiles y las estrellas de mar, menos voluminosas, tienen dos gónadas en cada brazo. Si bien se cree que los ancestros de los equinodermos modernos tenían una abertura genital, muchos organismos tienen múltiples gonoporos a través de los cuales se pueden liberar óvulos o espermatozoides.
Regeneración

Muchos equinodermos tienen grandes poderes de regeneración. Muchas especies autotomizan y regeneran rutinariamente brazos y vísceras. Los pepinos de mar a menudo descargan partes de sus órganos internos si se sienten amenazados, regenerándolos en el transcurso de varios meses. Los erizos de mar reemplazan constantemente las espinas perdidas por el daño, mientras que las estrellas de mar y los lirios de mar pierden y regeneran fácilmente sus brazos. En la mayoría de los casos, un solo brazo cortado no puede convertirse en una nueva estrella de mar en ausencia de al menos una parte del disco. Sin embargo, en algunas especies, un solo brazo puede sobrevivir y convertirse en un individuo completo, y los brazos a veces se separan intencionalmente con el propósito de la reproducción asexual. Durante los períodos en los que han perdido sus tractos digestivos, los pepinos de mar viven de los nutrientes almacenados y absorben la materia orgánica disuelta directamente del agua.
La regeneración de las partes perdidas implica tanto la epimorfosis como la morfalaxis. En la epimorfosis, las células madre, ya sea de un grupo de reserva o de aquellas producidas por desdiferenciación, forman un blastema y generan nuevos tejidos. La regeneración morfaláctica implica el movimiento y la remodelación de los tejidos existentes para reemplazar las partes perdidas. También se observa la transdiferenciación directa de un tipo de tejido a otro durante el reemplazo de tejido.
Reproducción
Reproducción sexual
Los equinodermos alcanzan la madurez sexual después de aproximadamente dos o tres años, según la especie y las condiciones ambientales. Casi todas las especies tienen sexos masculino y femenino separados, aunque algunas son hermafroditas. Los óvulos y los espermatozoides normalmente se liberan en aguas abiertas, donde tiene lugar la fertilización. La liberación de espermatozoides y óvulos está sincronizada en algunas especies, generalmente con respecto al ciclo lunar. En otras especies, los individuos pueden agruparse durante la temporada reproductiva, lo que aumenta la probabilidad de una fertilización exitosa. Se ha observado fertilización interna en tres especies de estrellas de mar, tres estrellas frágiles y un pepino de mar de aguas profundas. Incluso en las profundidades abisales, donde no penetra la luz, los equinodermos a menudo sincronizan su actividad reproductiva.
Algunos equinodermos empollan sus huevos. Esto es especialmente común en especies de agua fría donde las larvas planctónicas podrían no encontrar suficiente alimento. Estos huevos retenidos suelen ser pocos en número y se suministran con yemas grandes para nutrir a los embriones en desarrollo. En las estrellas de mar, la hembra puede llevar los huevos en bolsas especiales, debajo de los brazos, debajo de su cuerpo arqueado o incluso en su estómago cardíaco. Muchas estrellas frágiles son hermafroditas; a menudo incuban sus huevos, generalmente en cámaras especiales en sus superficies orales, pero a veces en el ovario o el celoma. En estas estrellas de mar y estrellas quebradizas, el desarrollo suele ser directo a la forma adulta, sin pasar por un estado larvario bilateral. Algunos erizos de mar y una especie de dólar de arena llevan sus huevos en cavidades, o cerca de su ano, manteniéndolos en su lugar con sus espinas. Algunos pepinos de mar usan sus tentáculos bucales para transferir sus huevos a la parte inferior o trasera, donde quedan retenidos. En un número muy reducido de especies, los huevos quedan retenidos en el celoma donde se desarrollan de forma vivípara, emergiendo posteriormente a través de rupturas en la pared del cuerpo. En algunos crinoideos, los embriones se desarrollan en bolsas de reproducción especiales, donde se guardan los óvulos hasta que el esperma liberado por un macho los encuentra.
Reproducción asexual

Una especie de estrella de mar, Ophidiaster granifer, se reproduce asexualmente por partenogénesis. En algunos otros asterozoos, los adultos se reproducen asexualmente hasta que maduran y luego se reproducen sexualmente. En la mayoría de estas especies, la reproducción asexual es por fisión transversal con el disco dividido en dos. Tanto el área del disco perdido como los brazos que faltan vuelven a crecer, por lo que un individuo puede tener brazos de diferentes longitudes. Durante el período de rebrote, tienen algunos brazos diminutos y un brazo grande, por lo que a menudo se les conoce como "cometas".
Los pepinos de mar adultos se reproducen asexualmente por fisión transversal. Holothuria parvula utiliza este método con frecuencia, dividiéndose en dos un poco antes del punto medio. Cada una de las dos mitades regenera sus órganos faltantes durante un período de varios meses, pero los órganos genitales faltantes suelen desarrollarse muy lentamente.
Las larvas de algunos equinodermos son capaces de reproducirse asexualmente. Se sabe desde hace mucho tiempo que esto ocurre entre las estrellas de mar y las estrellas frágiles, pero se ha observado más recientemente en un pepino de mar, un dólar de arena y un erizo de mar. Esto puede ser por autotomización de partes que se desarrollan en larvas secundarias, por gemación o por división transversal. Las partes o yemas autotomizadas pueden convertirse directamente en larvas completamente formadas, o pueden pasar a través de una gástrula o incluso una etapa de blástula. Las nuevas larvas pueden desarrollarse a partir de la capucha preoral (una estructura similar a un montículo sobre la boca), la pared lateral del cuerpo, los brazos posterolaterales o sus extremos traseros.
La clonación es costosa para la larva tanto en recursos como en tiempo de desarrollo. Las larvas se someten a este proceso cuando la comida es abundante o las condiciones de temperatura son óptimas. Puede ocurrir la clonación para hacer uso de los tejidos que normalmente se pierden durante la metamorfosis. Las larvas de algunos dólares de arena se clonan cuando detectan mucosidad de pescado disuelta, lo que indica la presencia de depredadores. La reproducción asexual produce muchas larvas más pequeñas que escapan mejor de los peces planctívoros, lo que implica que el mecanismo puede ser una adaptación anti-depredador.
Desarrollo larvario

El desarrollo comienza con un embrión bilateralmente simétrico, con una celoblastula que se desarrolla primero. La gastrulación marca la apertura de la "segunda boca" que coloca a los equinodermos dentro de los deuterostomas, y el mesodermo, que albergará el esqueleto, migra hacia adentro. La cavidad corporal secundaria, el celoma, se forma por la partición de tres cavidades corporales. Las larvas suelen ser planctónicas, pero en algunas especies los huevos quedan retenidos dentro de la hembra, mientras que en otras la hembra empolla las larvas.
Las larvas pasan por varios estadios, que tienen nombres específicos derivados de los nombres taxonómicos de los adultos o de su apariencia. Por ejemplo, un erizo de mar tiene un 'echinopluteus' larva mientras que una estrella quebradiza tiene un 'ophiopluteus' larva. Una estrella de mar tiene una 'bipinnaria' larva, que se convierte en una 'brachiolaria' larva. La larva de un pepino de mar es una 'auricularia' mientras que un crinoideo es una 'vitellaria'. Todas estas larvas son bilateralmente simétricas y tienen bandas de cilios con los que nadan; algunos, generalmente conocidos como 'pluteus' larvas, tienen brazos. Cuando están completamente desarrollados, se asientan en el lecho marino para sufrir una metamorfosis y los brazos y el intestino de las larvas se degeneran. El lado izquierdo de la larva se convierte en la superficie oral del juvenil, mientras que el lado derecho se convierte en la superficie aboral. En esta etapa se desarrolla la simetría pentarradial.
Se considera que una larva que se alimenta de plancton, que vive y se alimenta en la columna de agua, es el tipo de larva ancestral de los equinodermos, pero en los equinodermos existentes, alrededor del 68 % de las especies se desarrollan utilizando una larva que se alimenta de yema. La provisión de un saco vitelino significa que se produce una menor cantidad de huevos, las larvas tienen un período de desarrollo más corto y un menor potencial de dispersión, pero una mayor probabilidad de supervivencia.
Distribución y hábitat
Los equinodermos se distribuyen globalmente en casi todas las profundidades, latitudes y entornos del océano. Los adultos son principalmente bentónicos y viven en el lecho marino, mientras que las larvas suelen ser pelágicas y viven como plancton en el océano abierto. Sin embargo, algunos adultos holoturoides como Pelagothuria son pelágicos. Algunos crinoideos son pseudoplanctónicos y se adhieren a troncos y escombros flotantes, aunque este comportamiento se ejerció más ampliamente en el Paleozoico, antes de que la competencia de organismos como los percebes restringiera el alcance del comportamiento.
Modo de vida
Locomoción

Los equinodermos utilizan principalmente sus pies ambulacrales para moverse, aunque algunos erizos de mar también utilizan sus espinas. Los pies ambulacrales suelen tener una punta con forma de ventosa en la que se puede crear un vacío mediante la contracción de los músculos. Esto se combina con cierta pegajosidad de la secreción de moco para proporcionar adherencia. Los pies ambulacrales se contraen y relajan en ondas que se mueven a lo largo de la superficie adherente, y el animal avanza lentamente.
Las estrellas frágiles son las más ágiles de los equinodermos. Cualquiera de los brazos puede formar el eje de simetría, apuntando hacia adelante o hacia atrás. El animal entonces se mueve de forma coordinada, impulsado por los otros cuatro brazos. Durante la locomoción, los brazos propulsores pueden realizar movimientos de serpiente o de remo. Las estrellas de mar se mueven usando sus pies tubulares, manteniendo sus brazos casi inmóviles, incluso en géneros como Pycnopodia donde los brazos son flexibles. La superficie bucal está cubierta por miles de pies ambulacrales que se mueven a destiempo entre sí, pero no en un ritmo metacrónico; de alguna manera, sin embargo, los pies ambulacrales están coordinados, ya que el animal se desliza constantemente. Algunas estrellas de mar excavadoras tienen puntas en lugar de ventosas en sus pies ambulacrales y pueden "deslizarse" a través del fondo del mar a un ritmo más rápido.
Los erizos de mar usan sus pies ambulacrales para moverse de forma similar a las estrellas de mar. Algunos también usan sus espinas articuladas para empujarse o hacer palanca o levantar sus superficies orales del sustrato. Si un erizo de mar se vuelca, puede extender sus pies ambulacrales en un área ambulacral lo suficiente como para ponerlos al alcance del sustrato y luego unir sucesivamente los pies del área contigua hasta que se endereza. Algunas especies perforan la roca, generalmente triturando la superficie con sus piezas bucales.
.jpg)
Los pepinos de mar son generalmente animales lentos. Muchos pueden moverse en la superficie del lecho marino o excavar en la arena o el lodo mediante movimientos peristálticos; algunos tienen pies tubulares cortos en la superficie inferior con los que pueden arrastrarse a la manera de una estrella de mar. Algunas especies se arrastran usando sus tentáculos bucales, mientras que otras logran nadar con movimientos peristálticos o flexión rítmica. Muchos viven en grietas, huecos y madrigueras y apenas se mueven. Algunas especies de aguas profundas son pelágicas y pueden flotar en el agua con papilas palmeadas que forman velas o aletas.
La mayoría de los crinoideos son móviles, pero los lirios marinos son sésiles y se adhieren a sustratos duros mediante tallos. El movimiento en la mayoría de los lirios marinos se limita a doblar sus tallos, puede doblar y enrollar y desenrollar sus brazos; algunas especies pueden reubicarse en el lecho marino arrastrándose. Las plumas de mar son sueltas y suelen vivir en grietas, debajo de corales o dentro de esponjas siendo sus brazos la única parte visible. Algunas plumas de mar emergen por la noche y se posan en eminencias cercanas para aprovechar mejor las corrientes que transportan alimentos. Muchas especies pueden "caminar" por el fondo del mar, levantando el cuerpo con la ayuda de los brazos, o nadando con los brazos. Sin embargo, la mayoría de las especies de plumas de mar son en gran parte sedentarias y rara vez se alejan de su lugar de escondite elegido.
Alimentación
Los modos de alimentación varían mucho entre los diferentes taxones de equinodermos. Los crinoideos y algunas estrellas frágiles tienden a ser filtradores pasivos, atrapando partículas suspendidas del agua que pasa. La mayoría de los erizos de mar son herbívoros; los pepinos de mar se alimentan de depósitos; y la mayoría de las estrellas de mar son cazadores activos.
Los crinoideos atrapan las partículas de comida usando los pies ambulacrales en sus pínnulas extendidas, las mueven hacia los surcos ambulacrales, las envuelven en moco y las transportan a la boca usando los cilios que recubren los surcos. Los requisitos dietéticos exactos de los crinoideos se han investigado poco, pero en el laboratorio se pueden alimentar con diatomeas.
Las estrellas de canasta se alimentan en suspensión y levantan sus brazos ramificados para recolectar zooplancton, mientras que otras estrellas quebradizas usan varios métodos de alimentación. Algunos se alimentan en suspensión, asegurando partículas de comida con hebras de moco, espinas o pies ambulacrales en sus brazos levantados. Otros son carroñeros y comedores de detritos. Otros nuevamente son carnívoros voraces y capaces de atar a sus presas en el agua con un repentino cerco de sus brazos flexibles. Luego, las extremidades se doblan debajo del disco para transferir la comida a las mandíbulas y la boca.
Muchos erizos de mar se alimentan de algas, a menudo raspando la fina capa de algas que cubre la superficie de las rocas con sus piezas bucales especializadas conocidas como linterna de Aristóteles. Otras especies devoran organismos más pequeños, que pueden atrapar con sus pies ambulacrales. También pueden alimentarse de peces muertos y otras materias animales. Los dólares de arena pueden realizar una alimentación en suspensión y alimentarse de fitoplancton, detritos, piezas de algas y la capa bacteriana que rodea los granos de arena.
Los pepinos de mar a menudo se alimentan de depósitos móviles o de suspensión, utilizan sus podios bucales para capturar alimentos de forma activa y luego rellenan las partículas individualmente en sus cavidades bucales. Otros ingieren grandes cantidades de sedimentos, absorben la materia orgánica y pasan las partículas minerales no digeribles a través de sus intestinos. De esta manera, perturban y procesan grandes volúmenes de sustrato, a menudo dejando características crestas de sedimentos en el lecho marino. Algunos pepinos de mar viven infaunamente en madrigueras, con el extremo anterior hacia abajo y el ano en la superficie, tragando sedimentos y pasándolos a través de sus intestinos. Otros excavadores viven con el extremo anterior hacia arriba y esperan que los detritos caigan en las entradas de las madrigueras o rastrillen los escombros de la superficie cercana con sus podios bucales.
Casi todas las estrellas de mar se alimentan de detritos o son carnívoras, aunque algunas se alimentan de suspensión. Los peces pequeños que aterrizan en la superficie superior pueden ser capturados por pedicilarios y la materia animal muerta puede ser carroñera, pero las principales presas son invertebrados vivos, en su mayoría moluscos bivalvos. Para alimentarse de uno de ellos, la estrella de mar se desplaza sobre él, acopla sus patas tubulares y ejerce presión sobre las valvas arqueando el lomo. Cuando se forma un pequeño espacio entre las válvulas, la estrella de mar inserta parte de su estómago en la presa, excreta enzimas digestivas y licua lentamente las partes blandas del cuerpo. A medida que el músculo aductor del bivalvo se relaja, se inserta más estómago y cuando se completa la digestión, el estómago vuelve a su posición habitual en la estrella de mar con su harina de bivalvo ahora licuada dentro. Otras estrellas de mar evierten el estómago para alimentarse de esponjas, anémonas de mar, corales, detritos y películas de algas.
Defensa antidepredación
.jpg)
A pesar de su bajo valor nutricional y la abundancia de calcita no digerible, los equinodermos son presa de muchos organismos, incluidos peces óseos, tiburones, patos eider, gaviotas, cangrejos, moluscos gasterópodos, otros equinodermos, nutrias marinas, zorros árticos y humanos. Las estrellas de mar más grandes se alimentan de las más pequeñas; la gran cantidad de huevos y larvas que producen forman parte del zooplancton, consumido por muchas criaturas marinas. Los crinoideos, por otro lado, están relativamente libres de depredación.
Las defensas contra depredadores incluyen la presencia de espinas, toxinas (inherentes o liberadas a través de los pies ambulacrales) y la descarga de hilos pegajosos y enredadores por parte de los pepinos de mar. Aunque la mayoría de las espinas de los equinodermos son romas, las de la estrella de mar corona de espinas son largas y afiladas y pueden causar una dolorosa herida punzante ya que el epitelio que las cubre contiene una toxina. Debido a que atrapan el tejido conectivo, que puede cambiar rápidamente de un estado flácido a un estado rígido, los equinodermos son muy difíciles de desalojar de las grietas. Algunos pepinos de mar tienen un grupo de túbulos cuvierianos que pueden expulsarse como hilos largos y pegajosos de su ano para enredar y desactivar permanentemente a un atacante. Los pepinos de mar ocasionalmente se defienden rompiendo la pared de su cuerpo y descargando el intestino y los órganos internos. Las estrellas de mar y las estrellas frágiles pueden sufrir una autotomía cuando son atacadas, separando un brazo; esto puede distraer al depredador el tiempo suficiente para que el animal escape. Algunas especies de estrellas de mar pueden nadar para alejarse del peligro.
Ecología

Los equinodermos son numerosos invertebrados cuyos adultos juegan un papel importante en los ecosistemas bénticos, mientras que las larvas son un componente importante del plancton. Entre las funciones ecológicas de los adultos se encuentran el pastoreo de erizos de mar, el procesamiento de sedimentos de erizos de corazón y la alimentación en suspensión y depósito de crinoideos y pepinos de mar. Algunos erizos de mar pueden perforar rocas sólidas, desestabilizando las paredes rocosas y liberando nutrientes en el océano. Los arrecifes de coral también se perforan de esta manera, pero la tasa de acumulación de material carbonatado suele ser mayor que la erosión producida por el erizo de mar. Los equinodermos secuestran alrededor de 0,1 gigatoneladas de dióxido de carbono por año en forma de carbonato de calcio, lo que los convierte en importantes contribuyentes en el ciclo global del carbono.
Los equinodermos a veces tienen grandes cambios de población que pueden transformar los ecosistemas. En 1983, por ejemplo, la mortalidad masiva del erizo de mar tropical Diadema antillarum en el Caribe provocó un cambio de un sistema de arrecifes dominado por corales a uno dominado por algas. Los erizos de mar se encuentran entre los principales herbívoros de los arrecifes y suele haber un buen equilibrio entre los erizos y las algas marinas y otras algas en las que pastan. Una disminución en el número de depredadores (nutrias, langostas y peces) puede resultar en un aumento en el número de erizos, causando un sobrepastoreo de los bosques de algas marinas, lo que resulta en un "erizo de erizos" desnudo de algas. En la Gran Barrera de Coral, un aumento inexplicable en el número de estrellas de mar corona de espinas (Acanthaster planci), que pastan en el tejido de coral vivo, ha aumentado considerablemente la mortalidad de los corales y ha reducido la biodiversidad de los arrecifes de coral.
Base de datos de organismos modelo
Echinobase es la base de datos de organismos modelo de equinodermos con varios niveles de soporte para las siguientes especies: erizo de mar morado (Strongylocentrotus purpuratus), estrella murciélago (Patiria miniata), Erizo de mar verde jaspeado (Lytechinus variegatus), estrella de mar corona de espinas (Acanthaster planci), estrella de plumas (Anneissia japonica) y estrella de azúcar (Asterias rubens). El soporte incluye la integración del genoma con páginas de genes seleccionadas (para las primeras cuatro especies), así como capacidades de BLAST, JBrowse y descarga de datos. Echinobase también mantiene pautas de nomenclatura de genes, páginas comunitarias (investigadores, laboratorios, organizaciones) y la ontología anatómica de Echinobase.
Uso por humanos
Como alimento y medicina


En 2019 se cosecharon 129.000 toneladas de equinodermos. La mayoría de estos fueron pepinos de mar (158.000 toneladas) y erizos de mar (73.000 toneladas). Estos se utilizan principalmente para la alimentación, pero también en la medicina tradicional china. Los pepinos de mar se consideran un manjar en algunos países del sudeste asiático; como tales, están en peligro inminente de ser sobreexplotadas. Las especies populares incluyen el rodillo de piña Thelenota ananas (susuhan) y el pepino de mar rojo Holothuria edulis. Estas y otras especies se conocen coloquialmente como bêche de mer o trepang en China e Indonesia. Los pepinos de mar se hierven durante veinte minutos y luego se secan de forma natural y luego sobre un fuego que les da un sabor ahumado. En China se utilizan como base para sopas y guisos gelatinosos. Tanto las gónadas masculinas como las femeninas de los erizos de mar se consumen, particularmente en Japón y Francia. El sabor se describe como suave y derretido, como una mezcla de mariscos y frutas. Se han llevado a cabo ensayos de cría de erizos de mar para tratar de compensar la sobreexplotación.
En investigación
Debido a su robusto crecimiento larvario, los erizos de mar se utilizan ampliamente en la investigación, particularmente como organismos modelo en biología del desarrollo y ecotoxicología. Strongylocentrotus purpuratus y Arbacia punctulata se utilizan para este fin en estudios embriológicos. El gran tamaño y la transparencia de los óvulos permite la observación de los espermatozoides en el proceso de fecundación de los óvulos. El potencial de regeneración del brazo de las estrellas frágiles se está estudiando en relación con la comprensión y el tratamiento de enfermedades neurodegenerativas en humanos.
Otros usos
Las pruebas calcáreas o caparazones de los equinodermos son utilizados como fuente de cal por los agricultores en áreas donde la piedra caliza no está disponible y algunos se utilizan en la fabricación de harina de pescado. Cuatro mil toneladas de los animales se utilizan anualmente para estos fines. Este comercio a menudo se lleva a cabo junto con los criadores de mariscos, para quienes las estrellas de mar representan una gran amenaza al comerse su población cultivada. Otros usos de las estrellas de mar que recuperan incluyen la fabricación de alimentos para animales, el compostaje y la preparación de especímenes secos para el comercio de artes y oficios.
Textos citados
- Brusca, Richard C.; Moore, Wendy; Shuster, Stephen M. (2016). Invertebrados (3a edición). Sunderland, Massachusetts. ISBN 978-1-60535-375-3. OCLC 928750550.
- Dorit, R. L.; Walker, W. F.; Barnes, R. D. (1991). Zoología, edición internacional. Saunders College Publishing. ISBN 978-0-030504-7.
- Ruppert, Edward E.; Fox, Richard, S.; Barnes, Robert D. (2004). Zoología Invertebrada (7a edición). Cengage Learning. ISBN 81-315-0104-3.
Contenido relacionado
Infundíbulo
Hypericum perforatum
Reynoutria japónica