
Megafauna

In terrestrial zoology, the term megafauna (from Ancient Greek megas "great" + Latin fauna &# 34;animal") refers to "giant", "very large" or "big". The term is especially associated in current scientific literature with the late Pleistocene and Holocene megafauna that have been dying out in a process known as the Quaternary mass extinction from 50,000 years ago to the present. It thus includes large or gigantic animals that are considered archetypal of the last ice age, such as mammoths in the Northern Hemisphere and glyptodons in South America. The most commonly used threshold for including a taxon in the term megafauna is 44 kilograms or 100 kilograms. This therefore includes many species that are not popularly considered especially large, such as white-tailed deer and red kangaroos, and even humans, although the criteria is also used. more restrictive to apply it to those animals whose mass exceeds one metric ton (one million grams or megagram).
In practice, a common usage found in academic and popular writing refers to terrestrial animals larger than a human being and that are not (only) domesticated. It is also used to refer to the largest wild animals that exist, especially elephants, giraffes, hippos, rhinos, moose, condors, etc. Megafauna can be subcategorized by their trophic position into megaherbivores (eg deer), megacarnivores (lions), and more rarely, megaomnivores (bears).
Other common uses fall on giant aquatic species such as whales, any wild or domesticated land animals such as antelopes and bulls, and dinosaurs and other extinct giant reptiles.
The term is also applied to large (usually extinct) animals relatives of a more common or surviving type of animal, for example the 1 meter wingspan dragonfly of the Carboniferous period.
Ecological Strategy
Megafauna—in the sense of the largest birds and mammals—are generally K-strategist animals, with long lifespans, low population growth rates, low mortality rates, and few or no predators capable of killing a adult. These characteristics, although not exclusive to this megafauna, make them highly vulnerable to human overexploitation, which is believed to have led to mass extinction events on all continents except sub-Saharan Africa and southern Asia, the only places of the planet that today conserve an important part of the habitual terrestrial megafauna in most of the ecosystems before our expansion during the Pleistocene.
Evolution of large body size
An observation that has been made about the evolution of large body size says that the rapid rates of increase that are often observed over relatively short intervals of time are not sustainable over much longer periods. In an examination of mammalian body masses over time, the maximum possible increase in a given time interval found was a scale with the length of the interval raised to the power of 0.25. This is believed to reflect the occurrence, during a trend of increasing maximum body size, from a series of anatomical, physiological, environmental, genetic, and other constraints that must be overcome by evolutionary innovations before further increases in size are possible. A surprisingly rapid rate of change was found for large decreases in body mass, such as may be associated with the phenomenon of insular dwarfism. It was found that when generation length is normalized, the maximum rate for large reductions in body mass is 30 times greater than the maximum rate for large increases in body mass.
In land mammals

Following the Cretaceous-Tertiary mass extinction that wiped out the dinosaurs about 66 million years ago, land mammals experienced a near exponential increase in body size as they diversified to fill vacated ecological niches. Starting from just a few kilograms before the extinction, its maximum size increased to 50 kg a few million years later, reaching 750 kg in the late Paleocene. This trend of increasing body mass seems to stabilize around 40 million years ago (in the late Eocene), suggesting that ecological or physiological constraints had been reached, after a three-order-of-magnitude increase in body mass. However, if considered from the point of view of the rate of increase of the size of each generation, it is found that the exponential increase continued until the appearance of Paraceratherium 30 million years ago. (Given the generation time scales of body mass 0.259 , increasing generation times with increasing size cause the record of mass as a function of time to form a downward curve from a linear fit.)
Megaherbivores eventually attained a body mass of more than 10,000 kg. The largest of these, the indricoterians and proboscidians, were large intestinal fermenters, which are thought to have an advantage over small intestinal fermenters in terms of being able to speed up gastrointestinal transit to accommodate the ingestion of huge amounts of food. A similar trend appears when comparing rates of maximum body masses per generation for different mammalian clades (using rates averaged over macroevolutionary time scales). Among terrestrial mammals, the fastest rates of increase appear in perissodactyls, followed by rodents and proboscideans, all of which are large intestine fermenters. The rate of increase for artiodactyls was about one third that of perissodactyls. The rate for carnivores was slightly lower, while primates, perhaps restricted by their arboreal habits, had the lowest rate among the mammal groups studied.
The carnivorous land mammals of various groups of eutherians (the mesonychian Andrewsarchus, the creodonts Megistotherium and Sarkastodon, and the carnivorous Amphicyon and Arctodus) reached a maximum size of about 1,000 kg (apparently Arctotherium was somewhat larger). The largest metatherian carnivore, Proborhyaena gigantea, apparently reached 600 kg, also close to this limit. A theoretical maximum size limit has been predicted based on the metabolic rate of mammals, the energy cost of obtaining prey items, and the estimated maximum rate of prey consumption. It has also been suggested that the maximum size of carnivorous mammals is constrained by the stress that the humerus can withstand during maximum running speed.
Analysis of the variation in maximum body size over the last 40 million years suggests that falling temperatures and increasing continental land area are associated with increasing size. The first correlation could be consistent with Bergmann's rule, and could be related to the thermoregulatory advantage of greater body mass in cold climates, the better ability of large organisms to cope with food availability, or other factors; the latter correlation could be explainable in terms of range and resource constraints. However, the two parameters are interrelated (due to drops in sea level accompanying increased ice ages), making make the main reason for the size increase trends more difficult to identify.
In flightless birds
During the Paleocene, due to the small initial size of mammals, top predator niches were frequently occupied by members of other groups, such as terrestrial crocodilians (such as Pristichamsus), large snakes (Titanoboa), monitor lizards, or flightless birds (such as Gastornis in Europe and North America, Paleopsilopterus in South America). On the northern continents, large predatory birds were displaced when large eutherian carnivores evolved. In isolated South America, phorusrhacids could not be superseded by local sparasodont metatherian carnivores and remained dominant until eutherian predators arrived from North America (as part of the Great American Biotic Interchange) during the Pliocene. However, none of the larger land birds, such as the possibly omnivorous (Dromornis) or herbivorous (Brontornis and Aepyornis) of the Cenozoic grew larger beyond 500 kg, and therefore never reached the size of the largest mammalian predators, let alone the largest herbivorous mammals. It has been suggested that the increase in thickness of bird's egg shells in proportion to egg mass with increase in egg size implies an upper limit to the size of birds. The largest species of Dromornis, D. stirtoni, may have gone extinct after reaching maximum avian body mass and was later displaced by diprotodont marsupials which evolved to several times their sizes.
Late Quaternary mass extinctions
The Late Quaternary extinction event comprises all the megafauna mass extinctions recorded in the last 50,000 years, after the appearance of modern humans. These mass extinctions of megafauna began in the late Pleistocene, in tropical Asia and Oceania, continued in southern Europe between 40,000 and 20,000 years ago, peaked at the end of the last glacial period (the Würm glaciation) when the rest of the world died out. giant animals of the Holarctic region, such as the woolly mammoth, in the Americas and Eurasia, and later continued on islands and archipelagos around the world, the last major mass extinctions of megafauna being that of New Zealand between 900 and 500 years ago and the that are occurring in sub-Saharan Africa and tropical Asia today. The megafaunal extinctions known as the Holocene extinction and Pleistocene extinction today tend to fit by most scientists within the Late Quaternary extinction event, also known as the as QME (Quaternary Megafauna Extinction) for its acronym in English.
This megafaunal extinction more than halved the number of large mammals in the world, including large herbivores, predators, and scavengers. Australia lost 86% of its megafauna, most of it between 50,000 and 40,000 years ago, coinciding in time with the arrival of humans. In the case of both Americas, the extinction affected dozens of megafauna species, with a mass extinction that led to the disappearance of 73% of megafauna species in North America and 80% in South America, being the most widely accepted opinion today, that the American mass extinction also coincided with the arrival of humans in the New World. Thus, dating back at least 50,000 years and beginning in Asia and Australia, approximately 2/3 of all terrestrial mammal genera larger than 50 kilograms have become extinct in Eurasia, North Africa, Oceania, the Americas, and archipelagos around the world. World. Various theories have attempted to attribute the various waves of megafaunal extinctions of the past 50,000 years to non-anthropogenic causes such as climate change, unknown diseases (no known disease has caused the simultaneous extinction of totally different families, orders and even infraclasses, also discriminating species based on their size), a possible extraterrestrial impact (not recorded by geologists), among others.
Climate hypothesis
The climate hypothesis, which was dominant for most of the past 20th century, is unable to consistently explain these mass extinction patterns for several reasons:
- The climatic hypothesis does not explain the regional asynchrony of the exemptions. Climate events related to the last glacial cycles were planetary events that affected the entire globe at the same time: cold peaks for example were linked to dry periods in tropical regions. However, extinctions occurred in completely different periods in different parts of the world.
- There were more than 30 glacial cycles along Pleistocene at least as intense as the last, but without mass extinctions associated in the fossil record.
- Extinctions in southern Europe, tropical Asia, Oceania and almost all of the planet's islands did not coincide with glacial peaks or glacial cycle changes, but with climate-stable periods.
- The climatic hypothesis would involve many plant extinctions, which are often more affected by climate changes than animals, but this prediction fails ostensibly: there was no wave of massive floristic extinctions in the Quaternary.
- Extinctions present an unprecedented size selectivity in the fossil record, as they affect almost exclusively megafauna species, when the latest studies have revealed that they are the smallest and most vulnerable species and those that are extinct in greater proportion under environmental disturbances caused by climate.
- Fossils show in the period immediately prior to extinction a drastic reduction in the age of sexual maturity of megafauna species, which does not correspond to what is recorded in extinctions due to climatic factors, but has been studied and checked repeatedly in mammal populations subject to high kinetic pressure.
- Finally, climate change cannot explain why the extinctions have been systematically more recent on the islands than on the continents, when faced with climate change the expected is just the opposite: island populations, without the possibility of emigrating like the continental ones, and with a number of specimens and a much smaller genetic pool, should be extinguished first. However, the extinctions on the islands occurred already well into the Holocene, several thousand years after climate change (up to 5000 years ago in the case of the lazy giants in the Caribbean, 4000 years in that of the Siberian mammoths and the European elephants, and until the 18th in the case of the sea cow of Steller in the Bering Sea).
On the contrary, there does seem to be an increasingly clear correspondence between the dates of arrival of humans with those of the extinction of megafauna on each land mass.
Anthropogenic or overhunting hypothesis
The hypothesis of increasing acceptance among the scientific community is that these mass extinctions could be due fundamentally to the direct or indirect action of man due to an unsustainable level of exploitation of the megafauna. Until the beginning of the second decade of the 21st century, studies related to Quaternary extinctions had focused on certain continents or had used low-resolution analyzes and low-reliability dating. Statistical studies only began to be published in 2012. information on the causes of the Late Quaternary mass extinctions that included the latest advances in fossil dating and past climate models, thus developing the first high-resolution quantitative analyses, comparing the explanatory power of the hypotheses on a global scale. The statistical work published by Bernardo Araujo in 2015 applied mathematical models to compare the probability of influences of the s two variables (human arrival and climatic variation). This work offered as a result that the extinction phenomena were closer than expected due to chance, to the moment of human arrival in 65 cases (each case is the extinction of a gender in one of the 19 regions studied). On the other hand, the time of extinction was closer than expected by chance to periods of intense climate change in only 2 cases (a value close to that expected for a random ordering of extinctions over time). The extinctions were closer than expected due to chance both to human arrival and to times of intense climatic variation in 19 cases; and none of these variables in 39 cases, out of a total of 126 cases.
41 GLMM (Generalized Linear Mixed Model) models were also generated, covering a range of time intervals from 1,000 to 5,000 years. When either temperature proxy was used to represent climate variation for the entire planet, the 2H Antarctic detected a stronger climate effect than the 18O Greenlandic, but human arrival had an even stronger effect in both cases. Using each proxy to represent climate variation in its own hemisphere, the time of greatest climate variation was significantly related to megafauna extinction in only 1 of 41 intervals. In stark contrast, human arrival date was the best predictor of megafaunal extinction, with a significant effect at 40 of 41 intervals. In the models that considered the interaction between both factors, the interaction term was significant in only 4 of the 41 intervals. Thus, GLMM provided more evidence for anthropogenic than climatic effects, and little evidence for a synergistic action of the two factors.
These results provided clear support for the hypothesis that anthropogenic impacts, rather than climate variation, were the primary driver of the great Quaternary extinctions of megafauna.
Contrast between sub-Saharan megafauna and extinction in the rest of the planet: ecological naivety
All of the late Quaternary megafauna mass extinctions seem to have coincided in time with the expansion of modern humans to new continents or large islands, with sub-Saharan Africa being the least affected by the process until the 19th century. Today it is believed that sub-Saharan Africa was able to maintain its megafauna thanks to the fact that African species endured a long presence of hunting hominids since the appearance of the genus Australopithecus. Thus, these megafauna species were able to co-evolve with species of the genus Homo and other older genera, and develop an instinctive response to avoid contact with hominids or initiate an attack in their presence. The megafauna of the rest of the world or it had been exposed to a lesser presence of primitive humans although subjected to their predation (this is the particular case of the Eurosiatic / Palearctic megafauna, whose mass extinction was the slowest of all after the arrival of Homo sapiens), or it had never cohabited with hominids, so it would not have developed any instinctive mechanism to identify us as predators. This lack of defensive response is known as insular ingenuity or ecological ingenuity (island tameness or island naivety in English) and has been recorded as a factor of extinction on numerous occasions. It is currently observed in those endemic species of recently discovered islands and continents such as the Galapagos or Antarctica. This behavior is considered by ecologists as a key factor in the vulnerability of native species to the appearance of alien species. For example, the penguins of the Antarctic continent they show a deep instinct of evasion against predatory species with which they have co-evolved, such as the leopard seal, but not against humans, whom they do not fear.
The huge difference between the number of butchering points for African elephants and those used to butcher proboscideans hunted in other parts of the globe seems to support this hypothesis. Thus, there are more proboscis butchering sites in the United States than there are elephant butcher sites in all of Africa, a land mass that is much larger than the United States. Not only is the difference in size between Africa and the US remarkable, but hominin presence in Africa dates back at least 100 times as far back in time as the short chronostratigraphic span of human presence in North America. However, in Africa there are less than a dozen probable butchering sites, despite the fact that they cover a much wider range of coexistence times, from the Plio-Pleistocene to the present. In contrast, in the US there are at least 14 sites for a period of at least 100 times shorter coexistence. In Eurasia remains demonstrating intensive use of the woolly mammoth as a food source are even more abundant and give a idea of the extent to which this animal was hunted on a large scale by modern humans. There are more than 70 sites throughout the Russian plain, where not only individual houses have been found, but entire communities that were essentially built with mammoth bones, settlements where those bones were also used as fuel (and for this they had to be relatively fresh), and where there are huge traps dug into the permafrost, with remains of mammoth meat and bone remains.
Tropical Asia: an under-studied megafauna extinction
Although tropical Asia has traditionally been included as another region along with sub-Saharan Africa where there have been no megafauna mass extinctions, the truth is that it has also had very notable megafaunal losses that have been overlooked in many studies because they do not show a peak in the Late Pleistocene and many emblematic species have survived to the present day. The Asian mainland, all the large islands from Indonesia to New Guinea, and many more small islands, supported more large vertebrate species and much more megavertebrate biomass in the Pleistocene than they now harbor with many species seeing their original range minimized over the last 50,000 years—to 10% of their maximum range in the Holocene and further since the Pleistocene — or directly extinct such as the elephants Palaeoloxodon naumanni and Stegodon sp., the rhinos Rhinoceros philippinensis and Rhinoceros sinensis, the bread Asian giant golin, Asian aurochs, giant tapir Megatapirus augustus, or the New Guinea megamarsupials Hulitherium, Maokopia, Zygomaturus, and Protemnodon. This region is enduring an ongoing megafauna mass extinction episode that threatens surviving species.
Timeline of megafauna extinctions in Oceania, the Americas, and the large archipelagos
Outside the Afro-Eurasian mainland, where extinctions are more recent the further out of Africa, or from the landmass to the islands, these megafaunal extinctions also followed a distinctive land-mass-to-land-mass pattern, that followed human expansion into previously uninhabited regions of the globe, which as discussed shows no correlation with climate history (which can be visualized with graphs of recent geological time periods of climate markers such as marine oxygen isotopes or atmospheric carbon dioxide levels). Australia was first affected about 45,000 years ago, followed by Tasmania about 41,000 years ago (after the formation of a land bridge with Australia about 43,000 years ago), Japan apparently about 30,000 years ago, North America 13,000 years ago, South America about 500 years later, Cyprus 10,000 years ago, the West Indies 6,000 years ago, New Caledonia and nearby islands 3,000 years ago, Madagascar 2,000 years ago, New Zealand 700 years ago, the Mascarenas 400 years ago, and the Commander Islands 250 years ago. Almost all of the isolated islands of the The world could provide similar examples of extinctions that occurred shortly after the arrival of Homo sapiens, although many of these islands, such as Hawaii, never had a terrestrial megafauna, so their extinct fauna was smaller.
The extinction of megafauna in Oceania and its cause-effect relationship with environmental changes
An analysis of spores of the fungus Sporormiella (derived largely from megaherbivore excrement) in cores of bog sediments spanning the last 130,000 years from Lynch Crater in Queensland, Australia show that the megafauna had virtually disappeared around 41,000 years ago, at a time when climate change was minimal; the change was accompanied by the increase in charcoal of vegetable origin, and was followed by the transition from forests to fire-resistant sclerophyllous vegetation. The high resolution of the chronology of the changes supports the hypothesis that hunting by humans alone eliminated the megafauna, and that the subsequent change in the flora was most likely a consequence of the elimination of the browsers and the increase of the fires The increase in fires followed the disappearance of megafauna by nearly a century, and most likely resulted from the accumulation of combustible material when grazing stopped. In the following centuries the pastures increased; sclerophyll vegetation increased with an interval of another century, and sclerophyll forest developed after another thousand years. During two periods of climate change between 120 and 75 thousand years ago, sclerophyll vegetation had also increased at the site. in response to cooler, drier conditions; none of these episodes had a significant impact on the abundance of megafauna.
Mass extinctions of megafauna today
The great extinction pulse near the end of the Pleistocene and beginning of the Holocene was thus just one more in a series of megafauna mass extinctions that have occurred from 50,000 years ago to the present time in which the surviving megafauna of Africa and Asia, is also seeing its populations drastically reduced. Within the current mass extinction cycle we find ourselves in, in the period from 1970 to 2012, and according to a recent global analysis, there has been a general decline in 58% in the strength of vertebrate populations.
Mass extinctions before the spread of Homo sapiens
Several mass extinctions have occurred earlier in Earth's geological history, involving losses of megafauna. The best known is the Cretaceous-Tertiary boundary extinction in which most of the dinosaurs and several giant reptiles were wiped out. However, previous mass extinctions were global and not selective for megafauna; that is, countless species of other groups, including plants, invertebrates weighing less than 45 kilos, terrestrial and marine invertebrates, and plankton, also became extinct. Thus, earlier events must have been caused by more widespread disturbances in the biosphere than the irruption of a new local factor in each of the continent-wide and island-wide extinctions of the past 50,000 years.
Continued hunting and environmental disturbance by humans has led to a series of megafaunal extinctions in the recent past, and has created a serious risk of additional extinctions in the near future.
Consequences of the mass extinction of megafauna
The extinction of megafauna in a large part of Eurasia, in the Americas and in Oceania had devastating and catastrophic effects for terrestrial ecosystems, which continue to this day and whose dimensions in biodiversity and ecosystem losses are only beginning to be elucidate.
Effects on biodiversity and carrying capacity of ecosystems
With the extinction of megafauna, the relationship between different species changes, to the point that several plant species become extinct, and the surviving animals are affected in many ways. After the extinction of the megafauna, an increase in the accumulation of plant mass has been verified that eventually ends up causing large fires and severe changes in the relationship between species, their diversity and the carrying capacity of the ecosystem. Perhaps the most visible consequence today is the relative absence of large animals in relation to ecosystems that conserve the original megafauna, even in areas that are abandoned for thousands of years by humans after the extinction of the megafauna.
As an example, you can compare the numbers of national parks in North America without human presence today, and Africa. In Canada, Jasper National Park contains approximately 1,300 wapitis and 150 elk. The park is approximately 11,000 square km and is very well protected from poaching. By contrast, the similarly sized Chobe National Park in Botswana's Jasper suffers from poaching but supports among many other species more than 50,000 elephants.
The Serengeti is one of the least disturbed natural spaces in the world despite the increasing poaching it currently suffers. It is only 3 times the size of Jasper, but it contains 1.7 million wildebeest, 250,000 zebras and 500,000 gazelles. In addition to elephants, hippos, giraffes, rhinos, etc. In terms of predators, Jasper contains approximately 110 grizzly bears, 90 black bears, and only 45 wolves. The Serengeti has more than 3,000 lions, 8,000 hyenas, and 1,000 leopards. These numbers are even more remarkable when considering the increasing pressure of poaching on African wildlife in recent decades. The cape buffalo population in the Serengeti was 65,000 in 1969, but has dropped to 16,000 today.
Jasper has a shorter growing season, with less intense sunlight, while the Serengeti has suffered from poaching for four decades and severe seasonal drought. However, there is a huge difference in the number of animals over 44 kilos that live in both parks. According to the most recent studies, the fundamental reason for this huge difference in the size of the populations of large mammals would be the recent extinction (immediate on a geological scale) of the megafauna in North America caused by the irruption of Homo sapiens.
Today it is very difficult to estimate the number of animals that would be present if Jasper were still an untouched space. A study of fossil remains in Russia provides some figures for the subarctic grasslands of northern Siberia. The research findings suggest approximately 1 mammoth per square kilometer in one of the coldest environments on earth. It also provides figures for some other megafauna species. If the figures are extrapolated, Jasper could support about 11,000 mammoths, 66,000 horses and 165,000 deer. In Jasper, the role of European mammoths would likely be split between American mammoths and another huge elephant known as the mastodon. The area should also contain camels, giant sloths, giant beavers, American lions, jaguars, saber-tooth tigers, saiga antelopes, giant wolves, giant bears, American cheetahs, giant bison, various species of musk ox, and pronghorn.
Effects on nutrient transport
Megafauna play an essential role in the lateral transport of mineral nutrients in ecosystems, tending to relocate them from areas of great abundance to those of less quantity. This movement is due to the time between the consumption of the nutrients and the time in which they are released through excretion (or, to a lesser extent, through decomposition after death). It is estimated that in the Amazon Basin of South America, this lateral diffusion was reduced by 98% after the megafaunal extinctions that occurred approximately 12,500 years ago Since phosphorus availability is believed to limit productivity in most of the region, the decrease in this Transport from the western part of the basin and from the floodplains (both regions derive their supply from the Andean elevations to other areas) must have had a significant impact on the ecology of the region, and the effects may not have fully reached their full potential. limit.
Effects on seed dispersal
Some plants that depend on megafauna for seed dispersal are still alive and are slowly going extinct without it. A well-studied example is the Louisiana orange tree. This tree produces a large fruit that no American animal eats today. For the plant this is a huge loss of energy, since the purpose of any fruit is to disperse seeds through the animals that consume it. Since no modern animal eats it, the fruit falls to the ground and the seeds remain under the tree. In this way the range of distribution of these plants is contracted, instead of extended. When the Louisiana orange tree was first recorded by Europeans, its range was limited to a small territory in central/north Texas. However, in the fossil pollen record it appears widespread throughout the continent. In fact, today we know that there were 7 species, the other 6 decreased their populations to extinction without the megafauna. These large fruit trees should be much more common than they are today due to the extinction of megafauna.
In South America, a 2015 statistical study has revealed that dozens of tree species (more than 60) whose fruits are believed to be dispersed by megafauna, have seen their distribution drastically reduced since the extinction of these animals.
Effects of the disappearance of megafauna on methane emissions
Many herbivores produce methane as a byproduct of fermentation in digestion, and release it by belching. Large populations of herbivorous megafauna have the potential to greatly contribute to the atmospheric concentration of methane, which is an important greenhouse gas with a less long-acting but more intense action than CO2. Today, about 20% of annual methane emissions come from domestic livestock and are considered a key driver of the greenhouse effect. Recent studies have shown that the extinction of megafauna herbivores must have caused a drastic reduction in atmospheric methane. This hypothesis is relatively new but presents a series of strong arguments consistent with the effect described for current methane from livestock sources on global warming.
Several studies have examined the effect of removing megaherbivorous mammals on methane emissions. One of these studies examined methane emissions from bison that inhabited the North American Great Plains before the arrival of European settlers. This study estimated that the disappearance of bison caused a decrease of 2.2 Tg/yr. This is a proportionally very large change for this period of time in the planet's atmosphere, and is comparatively much smaller than what would have occurred after the extinction of half the megafauna during the last 50,000 years.
Thus, another study examined the change in the methane concentration in the atmosphere at the end of the Pleistocene, after the extinction of the megafauna of Eurasia and the American continent. After the first humans arrived in the Americas around 13,000 years before present, their unsustainable hunting and other associated ecological impacts led to the extinction of 73% of megafauna in North America and 80% in South America, by At the same time that huge populations of the woolly mammoth and other large palearctic mammals were disappearing in Eurasia. The results of the mathematical models suggest that this extinction produced a drop in methane production of ~9.6 teragrams per year. The records of ice cores at the poles support this hypothesis of the abrupt decrease in methane at this time. This suggests that the absence of methane emissions from megafauna may have caused the abrupt climatic cooling known as the Younger Dryas, a sudden climate change that authors supporting the climate change hypothesis had considered to cause the mass extinction of megafauna in the Americas and the Palaearctic, but in light of the latest research, it was probably a consequence of the same
Examples
The following are some notable examples of animals often considered megafauna (in the "large animal" sense). This list is not intended to be exhaustive:
- Class Mammalia
- Infraclase Metatheria
- Order Diprotodontia
- The red kangarooMacropus rufus) is the largest living mammal in Australia as well as the largest marsupial weighing 85 kg. However, his extinct relative, the short-faced kangaroo Procoptodon reached 230 kg of weight, while the extinct Diprotodon reached the largest size of any marsupial, estimated at more than 2,750 kg. The extinct marsupial lion (Thylacoleo carnifex), with more than 160 kg was much greater than any carnivorous marsupial. They became extinct after the arrival of the man.
- Order Diprotodontia
- Infraclase Eutheria
- Superorden Afrotheria
- Order Proboscidea
- The elephants are the greatest living terrestrial animals. These and their close relatives originated in Africa, but until recently they had an almost global distribution. The savannah African elephant (African Loxodonta) has a height to the shoulders of 4.3 meters and weigh more than 8 tons. Among the recently extinct mammoth tests (Mammuthus) were close relatives of the elephants, while the mastodons (Mammut) were much less related. They became extinct shortly after the man's arrival in America. It is estimated that the mammoth of the Songhua River (M. sungari) weighed up to 17 tons, making it one of the greatest proboscide and the second largest land animal after the indricoterins.
- Sirenia Order
- The largest modern sirenio with more than 1,500 kg weight is the manatí (Trichechus manatus). The sea cow of Steller (Hydrodamalis gigas) was probably five times larger, but fortunately was exterminated by humans in the 27 years following their rediscovery in the remote islands of the Commander in 1741. In prehistoric times this animal was also reproduced on the coasts of Northeast Asia and Northwest America; it was probably removed from these places most accessible by hunters.
- Order Proboscidea
- Superorden Xenarthra
- Order Cingulata
- The glypthodants were a group of large and heavily armored xenartros with a certain resemblance to the ankylosaurus dinosaurs and were relatives of the current armadillos. These appeared in South America, invaded North America during the Great American Biological Exchange, and became extinct at the end of Pleistocene.
- Order Pilosa
- The lazy land were another group of slow, earthly xenarms, related to the current lazy. These had a similar story, although they reached America long before, and expanded further north with Megalonyx. The greatest genders, Megatherium and EremotheriumThey reached sizes comparable to that of elephants. They became extinct after the man's arrival in America.
- Order Cingulata
- Superorden Euarchontoglires
- Order Primates
- The largest current primate, with more than 266 kg of weight, is gorilla (Gorilla beringei and Gorilla gorillawith three or four subspecies critically in danger. The extinct lémur lazy Malagasy, Archaeoindris reached a similar size, while the extinct is believed to be Gigantopithecus blacki Southeast Asia was several times greater. Some species of gender Homo were significantly higher than most of the current Homo sapiensfor example, Homo heidelbergensis In southern Africa it may have commonly reached 2.1 meters high, while the Neanderthals were about 30% more massive.
- Order Rodentia
- The current capibara (Hydrochoerus hydrochaeris) of South America, is the largest live rodent, weighing more than 65 kg. Several recently extinct American forms were greater: capibara Neochoerus pinckneyi (another neotropical migrant) that was 40% heavier; the giant castor (Castoroids ohioensis) was similar. The extinct giant jutia (Amblyrhiza flooded) of several Caribbean islands may have been even greater. However, several million years ago South America hosted much larger rodents. Phoberomys pattersoni, known of an almost complete skeleton, probably reached 700 kg. Fragmentary remains suggest that Josephoartigasia monesi it grew up to over 1,000 kg.
- Order Primates
- Superorden Laurasiatheria
- Order Carnivora
- Big felines include tiger (Panthera tigrisand the lion (Panthera leo). The largest subspecies, with more than 306 kg, is the Siberian tiger (P. tigris altaica), according to Bergmann's rule. Gender members Panthera are distinguished by morphological characteristics that allow them to roar. Among the largest extinct felines is the American lion (Panthera leo atrox) and the teeth of South American saber Smilodon populator. They became extinct shortly after the man's arrival in America.
- The bears are the biggest carnivores of the Caniformian suborden. The greatest living forms are the polar bear (Ursus maritimus), with a weight of more than 680 kg, and Kodiak bear (Ursus arctos middendorffi) of similar size, both also consistent with the Bergmann rule. Arctotherium angustidens, a short-sided South America bear, was one of the largest known land predatory mammals with an average weight of 1,600 kg.
- The pinípedos (morsas, focas and sea lions) are amphibian marine carnivores who evolved from ancestors similar to bears. The Marine Elephant of the South (Mirounga leonina) of the waters of the Antarctic and sub-antiartic is the largest of all carnivores, with adult males reaching a maximum length of 6 to 7 meters and a maximum weight of 5000 kilograms.
- Order Perissodactyla
- The tapirs are bouquetrous animals, with a prensil nose, something similar to a pig that seems to have changed little in the last 20 million years. These inhabit the tropical forests of Southeast Asia and Central and South America, and are among the largest existing land mammals in the last two regions.
- The rhinoceros are perisodactyls with horns made of keratin, the same type of protein is composed of the hair. These are among the largest living terrestrial mammals after the elephants (the hippopatamos reach a similar size). Three of the current five species are in critical danger. His former relatives of Central Asia were the largest land mammals of all time.
 Rinocerosant.
Rinocerosant.
- Order Artiodactyla (or classically, Cetartiodactyla)
- The giraffes (Giraffa camelopardalis) are the highest living terrestrial animals, reaching heights of about 6 meters.
- Bovine ungulped include the largest surviving animals in Europe and North America. The aquatic buffalo (Bubalus arnee), the bison (Bison bison and Bison bonasus) and the gaur (Bos gaurus) can grow to weights of more than 900 kg.
- The hippos (Hippopotamus amphibius) are the heaviest living artiodactyls; it is believed that this and the threatened dwarf hypopotamus (Choeropsis liberiensis) are close relatives of cetaceans. African hippopotisms are tracking the most dangerous megafauna species for humans.
- Order Cetacea (or classically, Cetartiodactyla)
- Cetaceans (ballenas, dolphins and marsopas) are marine mammals. The blue whale (Balaenoptera musculus) is the greatest of the rorcuals and the greatest animal that has ever lived. The cachalote (Physeter macrocephalus) is the largest dentist, as well as the most noisy animal with the greatest brain (which is five times larger than that of humans). The orca (Orcinus orca) is the largest dolphin.
- Order Carnivora
- Superorden Afrotheria
- Infraclase Metatheria
- Class Aves (phylogenetically, a nail inside Coelurosauria, a taxon within the Saurischia order of the Sauropsida clado; see later)
- Struthioniforms Order
- The ratites are an ancient and diverse group of non-flying birds found in the fragments of the former Gondwana supercontinent. The biggest bird today, the ostrich (Struthio camelus) was surpassed by the extinct Aepyornis of Madagascar, the heaviest of the group, and the extinct giant moa (Dinornis) of New Zealand, the highest, growing at heights of more than 3.4 meters. The last two are examples of insular gigantism.
- Anseriformes Order
- Australia's extinct drug addicts as Dromornis They may have exceeded the biggest rats in size. Because of its small size for a continent and its isolation, Australia is sometimes seen as the largest island in the world; therefore, these species could also be considered as island giants.
- Struthioniforms Order
- Class Reptilia (or classically, Sauropsida)
- Order Crocodilia
- The aligators and crocodiles are large semiaquatic reptiles, the largest of which, the saltwater crocodile (water crocodile)Crocodylus porosus), can grow to a weight of 1.360 kg. The distant ancestors of the crocodiles and their relatives, the crurotars, dominated the world at the end of the Triassic, until the extinction event of the Triassic-Jurassic allowed the dinosaurs to surpass them. These remained diverse at the end of the Mesozoic, when crocodiliforms as Deinosuchus and Sarcosuchus reached lengths of 12 m. Crocodilians of similar size, as Mourasuchus and PurussaurusThey existed as recently as the Miocene in South America.
- Order Saurischia
- The saurisquious dinosaurs of the Jurassic and Cretaceous include the sauropods, the longest terrestrial animals (with more than 40 meters) and the heavier ones known (Argentinosaurus reached between 80–100 metric tons, as well as the theropods, the largest terrestrial carnivores (Spinosaurus reached between 7 to 9 tons.
- Order Pterosauria
- The larger Azdaric pterosaurs, such as Hatzegopteryx and Quetzalcoatlus, they reached spans of about 11 to 12 m (36 to 39 feet) and weights probably in the range of 70 to 250 kg (150 to 550 lb). It is believed that the first was the main predator of its island ecosystem.
- Order Squamata
- Though the greatest living lizard, the dragon of Komodo (Varanus komodoensis), another island giant, can reach 3 meters long, its extinct Australian relative Megalania I could reach more than twice that size. The extinct marine relatives of the varanides, the mosaurids, were superdepredators in the seas of the Higher Cretaceous.
- The heaviest snake is considered to be the green anaconda (Eunectes murinus), while the reticulated python (Python reticulatus), with more than 8.7 meters, is the longest. An Australian extinct species of the Pliocene Liasis, the dubudingala, reached 10 meters, while Titanoboa the Paleocene of South America reached lengths of 12–15 meters and an estimated weight of about 1,135 kilograms.
- Order Testudines
- The greatest turtle is the threatened laúd turtle (Dermochelys coriaceaIt weighs more than 900 kg. It is distinguished from other sea turtles due to its lack of bone shell. The largest terrestrial quelonies are the giant turtles of the Galapagos Islands (Chelonoidis nigra) and the Aldabra atoll (Giant Aldabrachelys), with more than 300 kg. These turtles are the greatest survivors of a variety of giant turtle species that were widely represented in the continental land masses and several islands during Pleistocene.
- Order Crocodilia
- Class Amphibia
- Order Temnospondyli
- The Temnospondile of the Permic Prionosuchus, the greatest known amphibian, reached 9 meters long and was an aquatic predator similar to a crocodile. After the appearance of true crocodilians, fear them as Koolasuchus (from 5 meters long) they retired to the Antarctic region in the Cretaceous, before being extinguished.
- Order Temnospondyli
- Class Actinopterygii
- order Tetraodontiformes
- The greatest bonefish is the moonfish (Mola mola), whose average adult weight is 1,000 kg. Although phylogenetically it is a "pez bone", its skeleton is mostly cartilage (which is lighter than the bone). It has a disc-shaped body, and is driven with its long and thin dorsal and anal fins; it feeds mainly on jellyfish. In these three aspects (as well as their size and diving habits), it looks like the Laúd turtle.
- Acipenseriforms Order
- The threatened European sturgeon (Hustle) with more than 1476 kg) is the largest sturgeon (which are also mostly cartilageous) and is considered the largest anadrome fish.
- order Siluriformes
- The Mekong Cat Fish (Pangasianodon gigas), with more than 293 kg of weight, is often considered the largest freshwater fish.
- order Tetraodontiformes
- Class Chondrichthyes
- Lamniform Order
- The greatest live predator fish, the great white shark (Carcharodon carcharias), reaches up to 2,240 kg. Your extinct relative C. megalodon (discussion if it belongs to gender Carcharodon or Carcharocles) was greater by more than one order of magnitude, and is the greatest shark and predatory fish of any time; he hunted whales and other marine mammals.
- Orectolobiforms
- The largest current shark, cartilizing fish and fish in general is the whale shark (Rhincodon typus), which comes to weigh more than 21.5 tons. Like whales, it feeds by filtration and consumes mostly plankton.
- Rajiforms Order
- The stripe blanket (Manta birostris) is another species that feeds by filtration and is the largest strip, growing up to more than 2,300 kg.
- Lamniform Order
- Class Cephalopoda
- Order Teuthida
- A certain number of creatures of ocean depths exhibit abyssal gigantism. These include giant squid (Architeuthisand colossal squid (Mesonychoteuthis hamiltoni); it is believed that both cephalopodes (rarely seen) reach lengths of 12 meters or more. The last is the largest known invertebrate, and has the greatest eyes of any animal. Both are prey of the cachalotes.
- Order Teuthida

Gallery
Extinct
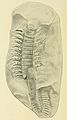
Fossil of a Arthropleura, a giant scolopendra that lived in the Carboniferous period.

Dunkleosteus It was a placoderm predator of the Devonian.

The pelicosaurus with candle Dimetrodon and the fear Eryops of the Permico of North America.

The plurid Pliosaurus (right) harassing the leaking fish Leedsichthys during the Jurassic.

Tyrannosaurus He was a theropode that was more than 12 m long, and he was the predator of western America.

Macronarian sauropods; from left to right, Camarasaurus, Brachiosaurus, Giraffatitan and Euhelopus.

The Earthling of Miocene Argentavis South America had a size of 8 meters.

The C. megalodon It's the biggest shark you know.

Deinotherium It had fangs turned down and was widely distributed in Afro-Eurasia.

Titanis walleri, the only bird of terror that is known invaded North America, measured 2 meters high.

The Diprotodon of Australia, the greatest marsupial of all time, was extinguished 40,000 years ago.

Megalania, a giant carnivorous rod from Australia, could have reached 4 meters long.

Mylodon, a type of giant lazy (sculpture located in the Osorno Pleistocenic Park.)

Megatherium, of the South American Pleistocene, was the size of an elephant and was the largest land lazy.

ToxodonOne of the biggest and last of the notungals. His relatives expanded to the United States.

The American lion exceeded the size of the present lion and lived in America until 10,000 years ago before the present.
![Los mamuts lanudos desaparecieron después de que los humanos invadieran su hábitat en Eurasia y América del Norte.[4]](https://upload.wikimedia.org/wikipedia/commons/thumb/a/a3/Woolly_mammoth_%28Mammuthus_primigenius%29_-_Mauricio_Ant%C3%B3n.jpg/120px-Woolly_mammoth_%28Mammuthus_primigenius%29_-_Mauricio_Ant%C3%B3n.jpg)
Naked mammoths disappeared after humans invaded their habitat in Eurasia and North America.

Haast eagle, the greatest known eagle, attacking a moa (which includes the highest known birds).









.jpg)

.jpg)



![Los mamuts lanudos desaparecieron después de que los humanos invadieran su hábitat en Eurasia y América del Norte.[4]](https://en.wikipedia.org/wiki/Archivo:Woolly_mammoth_(Mammuthus_primigenius)_-_Mauricio_Ant%C3%B3n.jpg)

Current species

The gorilla is the greatest primate and one of the most seriously threatened on the planet.

The Siberian tiger is the largest fertile, and an example of Bergmann's rule.

The polar bear is the largest bear and a semiaquatic carnivore, and are vulnerable to global warming.

The black rhinoceros, more than 4 meters long, is threatened by poaching.

Wild bactrian camels are seriously threatened. His ancestors originated in America.
![A diferencia del rinoceronte y el mamut lanudos, el buey almizclero logró sobrevivir a las extinciones del Cuaternario.[4]](https://upload.wikimedia.org/wikipedia/commons/thumb/e/ed/Ovibos_moschatus_qtl3.jpg/120px-Ovibos_moschatus_qtl3.jpg)
Unlike the rhinoceros and the lantern mammoth, the musk ox managed to survive the quaternary extinctions.

A hippopopotamus, the heaviest and most aquatic of artiodactyls, is the closest living relative of cetaceans.

The blue whale is a filter of more than 33 meters long and the greatest animal of all time.

The orca is the greatest dolphin and a hunter in herds, being very intelligent and social.

The ostrich eldest of the ratites, the heaviest bird, and, with up to 70 km/h, the fastest bird runner.

The saltwater crocodile is the greatest living reptile or danger to humans.

The dragon of Komodo, an island giant, is the largest lizard and has a dangerous infectious saliva.

The green anaconda, an aquatic constrictor, is the heaviest snake, with more than 97.5 kg.

Moonfish is the largest bone fish, but most of its skeleton is cartilizing.

The Nile Perch, one of the largest freshwater fish, is a dangerous invasive species.

Whale shark is the largest fish existing, growing up to more than 12.6 meters long.

The stripe blanket, a filtering species, is the largest of the stripes with more than 7.6 meters, although I could go back outside the water.

Review of a giant squid of 9 meters, an abyss giant and the second largest cephalopode.





![A diferencia del rinoceronte y el mamut lanudos, el buey almizclero logró sobrevivir a las extinciones del Cuaternario.[4]](https://en.wikipedia.org/wiki/Archivo:Ovibos_moschatus_qtl3.jpg)



.jpg)







Recommended bibliography
- Jared Diamond, 2006. Weapons, germs and steel. Editorial debate. ISBN 84-8306-667-X.
Contenido relacionado
Lepidoptera
Agenium
Elymandra