
Fossil

The fossils (from the Latin fosillis, which means 'excavated') are the remains or signs of the activity of past organisms. These remains, preserved in the sedimentary rocks, may have undergone transformations in their composition (by diagenesis) or deformations (by dynamic metamorphism) more or less intense. The science that deals with the study of fossils is paleontology. Within paleontology are paleobiology, which studies organisms from the past — paleobiological entities, which we know only by their fossil remains — biochronology, which studies when these organisms lived, and taphonomy, which deals with fossilization processes.
Etymology and evolution of the term



The word fossil is derived from the Latin verb fodere, to excavate, through the noun fossile, that which is excavated. Throughout history, and before, in prehistory, man has found fossils, remains of living beings petrified by the minerals with which they were in contact. It was those minerals that replaced or preserved its external form.
Primitive man attributed magical meaning to them. The authors of classical antiquity had observed them and, in general, correctly interpreted them. The term fossil was already used by Pliny in the 1st century, and its use was recovered in the 16th century by Agricola, alluding to its nature as a buried body (as a derivative of fossa) and included both organic remains and mineral bodies integrated into the materials of the earth's crust. This situation was maintained until the beginning of the last century, although it is true that the authentic fossils used to be differentiated as organized fossils.
The British geologist Lyell defined fossils as the remains of organisms that lived in other times and that are currently integrated within sedimentary rocks. This definition retains its validity, although currently the term has a broader scope, since it includes manifestations of the activity of organisms such as excrement (coprolites), remains of organic constructions, footprints, impressions of parts of the body, dentures (ichnofossils), etc.
Location

There are regions of the Earth that are known for their particular richness in fossils; for example, the Burgess Shale in British Columbia of Canada, the Solnhofen limestone, or the dinosaur-rich strata of Patagonia.
In Spain, Atapuerca and Las Hoyas stand out. The first is a rich Pleistocene site where, among other things, abundant hominid fossils have been found. The second is known for the presence of Iberomesornis.

Places that make exceptional preservation possible (sometimes even preserving soft tissue signs) are known as Lagerstätten (German for resting or storage places).
Types of Fossils
The oldest fossils are the stromatolites, which consist of rocks formed by the precipitation and fixation of calcium carbonate, thanks to bacterial activity. The latter has been known thanks to the study of current stromatolites, produced by microbial mats. The Gunflint Formation contains abundant microfossils widely accepted as microbial remains.
There are many kinds of fossils. The most common are remains of ammonoidea, snails or bones turned into stone. Many of them show all the original details of the shell or bone, even under a microscope. The pores and other small spaces in its structure fill with minerals.
Minerals are chemical compounds, such as calcite (calcium carbonate), that were dissolved in water. The passage through the sand or mud that contained the snails or the bones and the minerals were deposited in the spaces of its structure. That's why fossils are so heavy. Other fossils may have lost all marks of their original structure. For example, a snail shell originally made of calcite may completely dissolve after being buried. The impression left in the rock can be filled with other material and form an exact replica of the shell. In other cases, the shell dissolves and only the bone remains in the stone, a kind of mold that paleontologists can fill with plaster to discover the shape of the rest.
From a practical point of view we distinguish:
- nanofossil (visible under electronic microscope).
- microfossil (visible under optical microscope).
- macrofossil or megafossil (those we see in the naked eye).
Fossils usually show only the hard parts of the animal or plant: a tree trunk, a snail shell, or the bones of a dinosaur or fish. Some fossils are more complete: they record a greater amount of paleobiological information. If a plant or animal is buried in a special type of mud that does not contain oxygen, some of the soft parts may also become preserved as fossils.
The most spectacular of these "perfect fossils" they are complete woolly mammoths found in frozen ground. The meat was so frozen, it was still edible after 20,000 years. The most recent fossils, by convention, are those referring to organisms that lived at the end of the last Quaternary glaciation, that is, approximately 13,000 years ago. Later remains (Neolithic, Metal Age, etc.) are usually considered as subfossils.
Finally, those chemical substances included in the sediments that denote the existence of certain organisms that possessed or produced them exclusively must also be considered. They represent the extreme limit of the notion of fossil (biological markers or chemical fossils).
Ichnofossils

Ichnofossils are remains of depositions, footprints, eggs, nests, bioerosion or any other type of impression. They are the object of study of Paleoichnology.
Ichnofossils have their own characteristics that make them identifiable and allow their classification as parataxa: ichnogenera and ichnospecies. Parataxa are classes of fossil tracks grouped by their common properties: geometry, structure, size, substrate type, and functionality. Although sometimes diagnosing the species that produces an ichnofossil can be ambiguous, in general it is possible to infer at least the biological group or higher taxon to which it belonged.
Several types of behavior can be identified in ichnofossils: phyllotaxy, phobotaxis, helicotaxy, homostrophy, rheotaxy, thigmotaxis, and farming.
The term ichnofacies refers to the characteristic association of fossil tracks, recurring in space and time, that directly reflect environmental conditions such as bathymetry, salinity and the type of substrate. The tracks and footprints of marine invertebrates are excellent paleoecological indicators, as they are the result of the activity of certain organisms, related to specific environments, characterized by the nature of the substrate and conditions of the aquatic environment, salinity, temperature, and bathymetry. Especially the depth of the sea conditions the type of life of organisms and, therefore, it is not surprising that a whole series of ichnofacies can be distinguished according to bathymetry, whose nomenclature, due to Seilacher, refers to the type of most frequent tracks and more characteristics of each one.
An ichnofossil can have several interpretations:
- Philogenetics: Study the identity of the producer body. It gives way to the parataxons.
- Ethological: Study the behavior of the producer body.
- Taphono: It is interested in the original position and the taphonic processes suffered.
- Sedimentological: Reveal the paleoambient conditions of formation.
- Paleoecological: Studyed by icnophacies.
Microfossils

"Microfossil" It is a descriptive term that is applied when talking about those fossils of plants or animals whose size is smaller than that which can be analyzed by the human eye. Two diagnostic traits are commonly used to differentiate microfossils from eukaryotes and prokaryotes:
- Size: Eucariots are significantly larger in size to procarotes, at least mostly.
- Complexity of forms: The most complex forms are associated with eukaryotes, due to the possession of cytoskeleton.
Fossil resin
_rotated.JPG)
Amber (fossil resin) is a natural polymer found in many types of strata throughout the world, including in the Arctic. It is the fossilized resin of trees millions of years ago. It comes in the form of yellowish stones.
In amber you can find fossils of insects and other small animals that, at the time, were trapped by the resin.
Pseudofossil

Pseudofossils are visual patterns in rocks, produced by geological processes, that resemble the forms of living beings or their fossils; a classic example is pyrolusite (manganese oxide, MnO2) dendrites, which resemble plant debris. The misinterpretation of pseudofossils has generated certain controversies throughout the history of Paleontology. In 2003, a group of Spanish geologists questioned the organic origin of the Warrawoona fossils which, according to William Schopf, corresponded to cyanobacteria that constituted the first feature of life on Earth 3.5 billion years ago. The basis for such rethinking was that filamentous structures, similar to these putative Warrawoona microfossils, can be produced at room temperature and pressure by the combination, in an alkaline medium, of a barium salt and a silicate. A new study published in 2015 by the journal Proceedings of the National Academy of Sciences finally resolved the controversy. Researchers David Wacey and Martin Saunders used transmission electron microscopy to examine ultra-thin slices of microfossil candidates to build nanometer-scale maps of their size, shape, chemistry, and char distribution. This made it clear that the carbon distribution was unlike anything seen in authentic microfossils, revealing their mineral origin.
Living Fossil
A living fossil is an informal term used to refer to any living species that bears a strong resemblance to a species known from fossils (you could say that it is as if the fossil had "come to life").
Brachiopods are a perfect example of "living fossils". Lingula is a living brachiopod with fossils found throughout the Cenozoic. Another example is the coelacanth. It was a great surprise to find this fish off the coast of Africa in 1938, when it was thought that they had been extinct for 70 million years.
Fossil Record
The fossil record is the set of extant fossils. It is a small sample of the life of the past distorted and biased. It is not, moreover, a random sample. Any paleontological investigation must take these aspects into account, in order to understand what can be obtained through the use of fossils.
Representativeness of the fossil record

The total number of species (between plants and animals) described and classified amounts to 1.5 million. This number continues to increase, with approximately ten thousand insects being discovered every year (there is a great diversity of insects, 850,000 species are known). It is estimated that only a hundred bird species remain to be described (there is a low diversity of birds, since only 8,600 species are known). Estimates of possible living species are five million. About 300,000 fossil species are known, that is, 20% of the number of known living species and less than 6% of the probable ones. The fossil record spans from 3.77 billion years ago to the present; however, 99% of its representatives are found from 545 million years ago to now. These are staggering comparisons when we consider that the fossil record spans hundreds of millions of years and that living fauna and flora represent only an instant of geological time. If the conservation of fossils were acceptably good, it would be foreseeable that the number of extinct species would greatly exceed the number of current species.
There are several possible explanations for the relative poverty of extinct species:
- Strong growth in biodiversity over time. This causes experts to wonder if there was a lack of variety in the geological past.
- Since diversity is measured by the number of taxa (species, genres, families, etc.) that lived for a defined time interval, and that not all geological times have the same, we must take into account the fact that some parts of the geological column are better known than others. The number of paleontologists working in the Paleozoic and Precambrico represents a very low percentage; however, the extent of these lands is considerable.
- The most recent rocks bloom in larger areas because they are closer to the "high part of the lot"
Everything suggests that current diversity may not be appreciably higher than average for all the time since the Cambrian. Therefore the low number of extinct species cannot be satisfactorily explained by the idea that diversity increases with evolutionary progress. Species become extinct and are replaced by others over the course of geological time. A time frame of 12 million years has been suggested for a complete replacement of all species. The duration of the different biochrons is between 0.5 and 5 million years (2.75 million years the average biochron). Finally, as a conclusion, the number of extinct species estimated is:
(5× × 106)× × 545× × 1062,75× × 106=991× × 106{displaystyle (5times 10^{6})times {frac {545times 10^{6}}}{2,75times 10^{6}}}}}}=991times 10^{6}}}
Fossilization
For a bodily remains or a sign of an organism to be considered a fossil, a physical-chemical process must have occurred that affects it, known as fossilization. In this process, more or less profound transformations can be produced that can affect its composition and structure. This process is a function of time, so a certain interval must have elapsed from the moment of production of the rest for it to be considered a fossil. Fossilization is an exceptionally rare phenomenon, as most components of living things tend to decay rapidly after death.

Permineralization occurs after burial, when empty spaces in an organism (spaces that were filled with liquid or gas in life) are filled with groundwater, and the minerals it contains precipitate, filling those spaces.
In many cases the original remains of the organism have been completely dissolved or destroyed.
Decomposition processes
They are the main responsible in the world. Its effect is the rarity with which soft organic parts are preserved (0.01% of individuals in a marine community have only soft parts). The presence of soft parts are indicative of exceptional sedimentological and diagenetic conditions.
Aerobic decomposition processes
They are the fastest and most effective for biodegradation. Therefore, anoxic conditions are a prerequisite for the preservation of lightly mineralized and soft-part organisms. The oxygen demand for decomposition in an aerobic environment is very high (1 mol of Corg. requires 106 mol of O2). A standard reaction would be like this:
(CH2O)106(NH3)16H3PO4+106O2→ → 106CO2+16NH3+H3PO4+106H2O{displaystyle (CH_{2}O)_{106}(NH_{3})_{16}H_{3PO_{4}+106O_{2}rightarrow 106CO_{2}+16NH_{3}+H_{3}PO_{4}O_{2}O,!
Effects of decay
Decomposition is the main source of information loss in the fossil record, and mineralization is the only way to stop it. Tissues may be preserved as permineralizations, altered organic residues, or, with prolonged deterioration, as imprints. If the decomposition exceeds the mineralization, the tissues are destroyed and only refractories such as chitin, lignin or cellulose are preserved.
Decomposition characterization
Decay in the fossil record can be characterized at three levels:
- Identification of the breakdown and loss of information in the structure of fossil organisms.
- Recognition of particular minerals and geochemical markers associated with particular decomposition regimes.
- Preservation of fossil microbes involved in the decomposition process.
Origin, accumulation and preservation of organic matter

Most of it is recycled (giving rise to CO2) within the water column, particularly in the euphotic zone. A relatively small proportion of the organic matter produced becomes part of the adjacent sediments, and is affected by modifiers of organic flow (biostratinomics), which are photo-oxidation, microbial activity and detritivorous organisms.
Fossildiagenic processes
In addition to free lipids, organic matter includes biopolymers such as carbohydrates, proteins, chitin and lignin, some of which will be used for consumption and modification by benthic organisms and various microorganisms. The rest, not used in this way, can undergo polycondensation to form geopolymers, and becomes part of protokerogen, a precursor of kerogen. With the burial of the sediment, the increasing condensation and insolubilization produces the slow diagenetic conversion to kerogen that constitutes the volume of organic matter in ancient sediments.
Biological markers and their uses
Organic molecules (chemical fossils) are abundant in many sediments and sedimentary rocks, and are called biomarkers. Their study and identification require sophisticated sampling and analysis techniques. They keep a very detailed record of past biological activity and are related to current organic molecules. The possible sources of biomarkers in geological samples are as many as there are known molecules in organisms.
Source rocks in the generation of hydrocarbons
A source rock is a rocky volume that has generated or has been generating and expelling hydrocarbons in sufficient quantities to form accumulations of oil and gas. Most potential source rocks contain between 0.8 and 2% organic carbon. An approximate limit of 0.4% is accepted as the lowest volume of organic carbon for the generation of hydrocarbons, with the optimum being above 5-10%. The nature of the hydrocarbons generated depends fundamentally on the composition of the kerogen, which can be made up of two types of organic matter:
- Provider of remains of terrestrial plantsin which case the sediments will release gas mainly.
- Provider of aquatic media (marine or lacustre) with bacteria, algae, phyto and zooplankton, in which case they will produce oil with sufficient ripening.
Destructive physical-chemical processes

The durability of skeletons is their relative resistance to fracture and destruction by physical, chemical and biotic agents. These destructive processes can be divided into five categories that follow a more or less sequential order:
- Disarticulation: It is the disintegration of skeletons made up of multiple elements along pre-existing joints or joints (it can occur even before death, as in mute or exuvios of many arthropods). Decomposition destroys the ligaments that bind the oses of equinodermos in a few hours or days after death. The ligaments such as those of the bivalves, composed of conkyolin, are more resistant and can remain intact for months despite the fragmentation of the shells.
- Fragmentation: It is produced by the physical impact of objects and by biotic agents such as predators (even before death) and scavengers. Some forms of breakage allow us to identify the predator. The shells tend to break along pre-existing weakness lines such as growth lines or ornamentation. The resistance to fragmentation is based on several factors:
- Morphology of skeleton.
- Membership.
- Microstructure, thickness and percentage of organic matrix.
- Abrasion: It is the result of polishing and grinding of skeletal elements, producing rounding and a loss of surface details. Semi-quantitative studies of abrasion proportions have been conducted, introducing shells into a rotating drum with siliceous gravel. Its degree of intensity is related to various factors:
- The energy of the medium.
- The time of the exhibition.
- The size of the abrasive agent particle.
- The microstructure of skeletons.
- Bioerosion: It can only be identified when associated with recognizable fossils such as clion sponges and endolytic algae. Its destructive action is very high in shallow marine environments, where you can now see a weight loss of 16 to 20% in contemporary shells. It is unclear whether these proportions were kept in the Paleozoic, when the clion sponges were less abundant.
- Corrosion and dissolution: It is the result of chemical instability of minerals that form skeletons in the water column or in the sediment pores. Dissolution can start at sediment-water interphase and can continue to considerable depths within the sediment. The bioturbation of the sediments normally favors the dissolution by the introduction of marine water within the sediment that at the same time favors the oxidation of sulphides.
- Corrasion: In practice, the effects of mechanical abrasion, most of those of bioerosion and corrosion are difficult to distinguish in fossils. Some authors propose the term of corrasion to indicate the general state of shells, resulting from any combination of these processes. The degree of corrasion provides a general time index that the remains have been exposed to these three processes.
The destructive processes of disarticulation, fragmentation and corrosion are very evident in the fossil record. These processes affect different types of skeletons differently. Most marine organisms can be assigned to one of five skeletal architectural categories: solid, arborescent, univalve, bivalve, or multi-element.
- Solid skeletons: Resistant to breakage and very resistant to mechanical destruction. However, by remaining on the sea floor prolonged intervals of time, they often have corrasion effects in greater magnitude than other skeletons.
- Arborescent skeletons: They are the most sensitive indicators of fragmentation; an absence of breakage in such skeletons is an excellent indicator of minimal sedimentary disturbance.
- Bivalve skeletons: They disarm relatively quickly after death, although those with conkyolin ligaments may remain articulated for prolonged periods.
- Multiple elements skeletons: They are the best indicators of rapid burial.
When the different types of skeletons and their sensitivities to destructive agents are taken together, we find some excellent indicators of sedimentary processes, which can be used to define different taphofacies.
Transportation and hydrodynamics
If the skeletal remains of organisms are considered as sedimentary particles, studies can be carried out on their hydrodynamic behavior (shells of brachiopods, bivalves, gastropods, cephalopods, ostracods and crinoids). In general, little is known about the hydrodynamic behavior of these hard parts, which are so abundant and ecologically important in shallow water environments from modern environments and from the fossil record. The hydrodynamic behavior of shells is complex and unpredictable, mainly due to the great diversity of shapes involved.
Fossildiagenesis

Understanding the diagenetic processes is essential to correctly interpret the original mineralogy, structure of skeletons and shells, their taxonomic affinities and their paleoecology. A problem that arises very frequently is to deduce what has been the original mineralogy of extinct groups (rugose corals, archaeocyatids, stromatoporids...). The transition to the fossil state is highly dependent on skeletal composition.
Carbonated Skeletons
After burial, carbonate is altered to a greater or lesser extent during early diagenesis.
Aragonite Skeletons
Aragonite is normally transformed into calcite through one of these main processes:
- Total dissolution: If the waters of the empty zone are not saturated in carbonates, the total dissolution of the skeleton and the filling by calcite is produced. The empty area reproduces a shell mould and the shell structure is not preserved. Druze can be formed with crystals towards the center. The time that lasts the process is variable.
- Calcification: In this second case the skeletons of the shells preserve the relict structures (other layers or lamelas of the shells). You can even preserve whole Aragonite crystals that give us valuable information. The replacement occurs gradually and respects the original structure.
Calcite skeletons
In general, fossil skeletons that were made of calcite frequently maintain this original composition (unless they have been silicified or dolomitized). The magnesium content tends to be reduced, so that there may be diagenic alteration with high or low calcite content. There are special techniques such as cathodoluminescence that make it possible to determine their original content from relict areas that have preserved their original composition.
Carbonate nodules and lithographic limestone
The preservation of soft parts is often associated with the precipitation of carbonates in the form of nodules and layers, as is the case of lithographic limestones. Carbonate nodules are made up of siderite or calcite and associated with clayey sediments rich in microorganisms. They contain fossils that are often preserved in three dimensions, sometimes including fossilized soft parts. Its size varies between 10 and 30 centimeters although they have been found up to 10 meters (including a complete plesiosaur). The content of microorganisms and their decomposition are the primary factors that control the degree of anoxia, Eh and pH. In the presence of oxygen, microbial respiration produces CO2 that accumulates in the pore water of the sediment, favoring the dissolution of carbonates.
H2O+CaCO3+CO2 2HCO3− − +Ca2+{displaystyle H_{2}O+CaCO_{3}+CO_{2}Longleftrightarrow 2HCO_{3}^{-}+Ca^{2+}}
In the absence of oxygen, the sediment bacteria use a series of alternative oxidants in the respiration process (Mn, NO3-, Fe or SO42-) and when all the oxidants have disappeared, it is the fermentation reactions that dominate, taking place methane. Lithographic limestones are formed in lacustrine and marine environments, they are very fine-grained and finely banded. An example is the famous Solnhofen limestones from the Jurassic of Bavaria which contain the fossils of Archaeopteryx. The carbonate in these deposits can originate from a biogenic source (such as calcareous algae) or as a chemical precipitate.
Pyritized Fossils
Sedimentary pyrite occurs as a minor component of both present and ancient clastic marine sediments. Current sediment studies have shown that authigenic pyrite formation usually occurs in very early diagenesis just a few centimeters below the water-sediment interface. An increase in the number of microorganisms or the depth of burial prevents the diffusion of oxygen into the sediment and the microorganisms are forced to respire anaerobically. Mineralization stops the loss of information associated with the decomposition of macroorganisms and the precipitation of pyrite in early diagenesis is an important means for the preservation of fossils. In soft tissues such as muscle and also chitin, pyritization may occur during early diagenesis. When the decomposition is more advanced and therefore the formation of pyrite later, soft tissues will be destroyed and only resistant biological compounds (called refractories) such as cellulose and lignin are preserved. Hard biogenic parts such as shells (calcium carbonate and magnesium) and bones (calcium phosphate) are some of the most decay-resistant biological structures. Of the two, calcium carbonate is the more unstable and therefore the one most likely to be replaced by pyrite. Sedimentary pyrite presents several morphologies:
- Framboids: Add spherical microcrystals in the form of cubes and octagons in clay sediments. Its size varies from microns to about 1 millimeter in diameter.
- Pyritized segments: They are sediments infiltrated into biogenic cavities that have been consolidated by pirita. They can replace the detritic grains.
- Filling of cavities: The euhedral stick of cavities is very common in clay sediments. These cavities in many cases constitute the space occupied by mollusks, brachypods and bones.
- Embedded: It is precipitation on the outer surface of fossils.
- Pseudomorphic textures: In pseudomorphosis, the pirita can replace both detritic and fossil minerals, including also the preservation of sedimentary structures, holes and coprolites.
Pyrite formation is controlled by the concentration of organic carbon, sulfate, and iron detrital minerals. In a normal marine environment iron minerals and sulphates are present in abundance and pyrite formation is controlled by the supply of organic carbon. However, in freshwater environments the formation of pyrite is very limited by the low concentration of sulfate.
Fossil preservation as primary phosphate
Phosphorus is a fundamental element in life. It is concentrated in hard tissues, such as bones or some cuticles, or more often in soft tissues. It is therefore not surprising that it is involved in fossilization. The skeleton of vertebrates is mainly composed of hydroxyapatite (Ca10(PO4)6(OH)2). Some OH can be locally replaced by F- ions, especially in teeth, producing a less soluble hydroxy-fluoroapatite. Invertebrate phosphate shells have similar compositions with some variation. The composition of fossil bones contain more fluorine. The mean fluoride volume of freshwater and marine fish bones is respectively 4,300 ppm and 300 ppm, while fossils contain 22,100 ppm and 19,900 ppm fluoride.
Calcareous Skeletons
Calcium carbonate skeletons can transition to apatite with no change in external morphology. In natural environments, this diagenic alteration is associated with phosphate deposits. The bacterial transformation of calcareous organisms on apatite has been demonstrated in the laboratory. These observations and experiments suggest the following possible mechanism:
- The phosphorus needed to replace carbonate by apatito comes from the sediment microorganisms.
- Micro-organisms (bacteria, algae, fungi) promote decomposition, freeing phosphate ions and acidifying the interstitial water of sediments (this acidification that can be very localized, promotes the dissolution of carbonates). The released phosphate is combined with calcium to form apatito that forms preferably in the carbon/microorganism interface replacing the dissolved carbonate. This replacement preserves the external shape of the original shell and as in the fossilization of the primary apatito, the fluoride plays an important role as the final carbonate-fluor-apatito composition.
Siliceous skeletons
Primary silica phosphatization also appears in some radiolarian skeletons although this process is not yet well understood.
Microscopic examination of phosphorite samples shows that numerous microorganisms without a mineralized shell (algae, fungi, bacteria) mineralize as apatite, even though they did not have any mineral precursor. A well-known example is phosphatized coprolites where the organic matter itself is replaced by apatite that preserves the exact shape of the object. For example, the contraction striations of some coprolites. Soft tissue phosphatization is also frequent; many examples are known in arthropods (copepods, ostracods) that appear in calcareous and phosphate nodules within nodular limestone, or in coprolites of large vertebrates.
Studies on phosphorites and on the experimental synthesis of apatite have made it possible to estimate the probable conditions in the fossilization of apatite. Due to its stability requirements, apatite preferentially forms in an oxygen-deficient environment, sometimes even under fully reducing conditions, as indicated by its frequent association with pyrite. This environment is easily achieved in environments with abundant organic matter, which is in turn the main source of phosphorus.
Silica can replace calcite and aragonite in shells and permineralize wood. It can also form flint nodules or layers, replacing carbonate sediments or directly precipitating, enveloping or filling fossils or even remains of bacteria, organic microfossils and plants that are exceptionally preserved, as in the Rhynie Chert (Scotland).
There are three common modes of shell mineral replacement:
- Like a granular white cortex.
- As a finely granular replacement.
- Like concentric silica rings.

Fossilization of plant remains
Plants are composed of several parts (stem, branches, roots, leaves, pollen, fruit, seeds) some of which separate during life, while others do so after death. An adequate understanding of the dispersal processes that affect these parts is very important in the correct interpretation of paleofloristic associations. Studies on leaf dispersal by wind show that it is determined by its weight and shape.
Vegetable remains can be preserved in various ways:
- Permineralization.
- Preservation of the original material.
- Carbonization.
DNA in fossils
Recently, the possibility of extracting DNA remains from fossils and amplifying them using PCR has been confirmed. The evolution of this knowledge has been very rapid, since if at the end of the 90s there were doubts about the veracity of the DNA fossil remains, by the year 2000 there were already publications and a methodology had been established. By then already short sequences of Neanderthal and mammoth fossils had been extracted. Years later, there are also many examples in plants and even bacteria. Thus, Golenberg and his team obtained a partial chloroplast DNA sequence belonging to a Magnolia latahensis fossil. has maintained controversy about the reliability of the procedures used. This fossil DNA would allow establishing phylogenetic relationships between different taxa, in addition to facilitating a global vision of the evolutionary branches. In addition, it facilitates the estimation of the existing mutation rate between related taxa. The proposed methods are:

- Amber extraction: This suggestion, in an inviable and fictitious principle, was fed into popular fantasy through fiction novel (and later film) Jurassic Park. In this book it was suggested that insects suckers trapped in amber could magnificently preserve DNA from other animals, such as dinosaurs. In reality it has been possible to extract DNA from insects preserved in amber of an antiquity greater than 100 million years, however the DNA fragments thus obtained so far correspond to the insects themselves, not to other animals that could have been fed.
- Extraction of crystals in bones: It has been observed that in the bones sometimes crystals form. Scientists showed that the DNA contained in these crystals was preserved in a relative good state.
- Direct removal of fossil: Argentinian scientists ensure that DNA stays even millions of years, so they are found directly in the remains.
Scientific significance

Fossils are of considerable importance for other disciplines, such as Geology or Evolutionary Biology, they are the practical applications of Paleontology.
Based on the succession and evolution of the species in the course of geological times, the presence of fossils allows us to date the layers of the terrain (Biostratigraphy and Biochronology), with greater or lesser precision depending on the taxonomic group and degree of conservation. This is how most of the divisions and units of the chronological scales used in stratigraphy have been established.
They provide information on paleobiogeographic, paleoclimatic, sedimentary paleoenvironments, the diagenetic evolution of the rocks that contain them, etc.
The fossils continue to be revised, each time using more modern techniques. The application of these techniques entails new observations that sometimes modify previous approaches. Thus, for example, after a review carried out in 2006 with X-ray tomographic techniques, it was concluded that the family that contains the Markuelia worms had a great affinity with the priapulid worms, and is adjacent to the evolutionary branch of Priapulida, Nematoda and Arthropoda.
Image gallery

Three small amonite fossils, each of about 1.5 cm in diameter

Eocene fossil fish, Priscacara liopsof the Green River formation of Wyoming

Asaphus kowalewskii, a permineralized trilobit

Megalodon teeth and Carcharodontosaurus. The latter was found in the Sahara Desert

Marine fish and crustaceans of Cenomaniense (Cretaceous) Lebanon

Petrified Forest National Park Tronco, Arizona, United States

Cone petrified Araucaria mirabilis of Patagonia, Argentina of the Jurassic period (210 Ma approx.)

Pliocene gastropode fossil of Cyprus with the tube of a serpulite worm attached

fossil Orthoceras of the Silurico

Fossil of an Eocene Flower, Florissant, Colorado, United States

A fossil of a sea urchin, Micraster, known as "Hadas bread" in the UK

Ventral Valve of a Roadiense Brachypode (Guadalupiense, Middle Permico) by Glass Mountains, Texas, United States
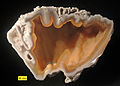
Coral (Oligocene-Miocene), Florida, United States. An example of preservation by agate substitution

Fossils of the beaches of Gotland Island, in the Baltic Sea (scores of 7 mm)













Contenido relacionado
Arctic Ocean
Kaolinite
Volcanic caldera