
Zona de brodmann
Un área de Brodmann es una región de la corteza cerebral, en el cerebro humano o de otros primates, definida por su citoarquitectura, o estructura histológica y organización de las células. El concepto fue introducido por primera vez por el anatomista alemán Korbinian Brodmann a principios del siglo XX. Brodmann cartografió el cerebro humano basándose en la variada estructura celular de la corteza e identificó 52 regiones distintas, que numeró del 1 al 52. Estas regiones, o áreas de Brodmann, se corresponden con diversas funciones que incluyen sensación, control motor y cognición.
Historia

Las áreas de Brodmann fueron definidas y numeradas originalmente por el anatomista alemán Korbinian Brodmann basándose en la organización citoarquitectónica de las neuronas que observó en la corteza cerebral utilizando el método de tinción celular de Nissl. Brodmann publicó sus mapas de áreas corticales en humanos, monos y otras especies en 1909, junto con muchos otros hallazgos y observaciones sobre los tipos celulares generales y la organización laminar de la corteza de los mamíferos. El mismo número de área de Brodmann en diferentes especies no indica necesariamente áreas homólogas. Constantin von Economo y Georg N. Koskinas publicaron un mapa cortical similar, pero más detallado, en 1925.
Importancia actual
Las áreas de Brodmann han sido discutidas, debatidas, refinadas y renombradas exhaustivamente durante casi un siglo y siguen siendo la organización citoarquitectónica de la corteza humana más conocida y citada con frecuencia.
Desde entonces, muchas de las áreas que Brodmann definió basándose únicamente en su organización neuronal se han correlacionado estrechamente con diversas funciones corticales. Por ejemplo, las áreas 1, 2 y 3 de Brodmann son la corteza somatosensorial primaria; el área 4 es la corteza motora primaria; el área 17 es la corteza visual primaria; y las áreas 41 y 42 corresponden estrechamente a la corteza auditiva primaria. Las funciones de orden superior de las áreas corticales de asociación también se localizan consistentemente en las mismas áreas de Brodmann mediante imágenes neurofisiológicas, funcionales y otros métodos (por ejemplo, la localización consistente del área del habla y el lenguaje de Broca en las áreas izquierdas de Brodmann 44 y 45).). Sin embargo, las imágenes funcionales sólo pueden identificar la localización aproximada de las activaciones cerebrales en términos de áreas de Brodmann, ya que sus límites reales en cualquier cerebro individual requieren su examen histológico.
Descripción general

Diferentes partes de la corteza cerebral están involucradas en diferentes funciones cognitivas y conductuales. Las diferencias se manifiestan de varias maneras: los efectos del daño cerebral localizado, los patrones de actividad regional expuestos cuando se examina el cerebro mediante técnicas de imágenes funcionales, la conectividad con áreas subcorticales y las diferencias regionales en la arquitectura celular de la corteza. Los neurocientíficos describen que la mayor parte de la corteza (la parte que llaman neocorteza) tiene seis capas, pero no todas las capas son evidentes en todas las áreas, e incluso cuando hay una capa presente, su grosor y organización celular pueden variar. Los científicos han construido mapas de áreas corticales basándose en las variaciones en la apariencia de las capas vistas al microscopio. Uno de los esquemas más utilizados provino de Korbinian Brodmann, quien dividió la corteza en 52 áreas diferentes y asignó a cada una un número (muchas de estas áreas de Brodmann se han subdividido desde entonces). Por ejemplo, el área 1 de Brodmann es la corteza somatosensorial primaria, el área 17 de Brodmann es la corteza visual primaria y el área 25 de Brodmann es la corteza cingulada anterior.

Muchas de las áreas del cerebro definidas por Brodmann tienen sus propias estructuras internas complejas. En varios casos, las áreas del cerebro están organizadas en mapas topográficos, donde partes contiguas de la corteza corresponden a partes contiguas del cuerpo, o de alguna entidad más abstracta. Un ejemplo sencillo de este tipo de correspondencia es la corteza motora primaria, una franja de tejido que se extiende a lo largo del borde anterior del surco central. Las áreas motoras que inervan cada parte del cuerpo surgen de una zona distinta, con las partes del cuerpo vecinas representadas por zonas vecinas. La estimulación eléctrica de la corteza en cualquier punto provoca una contracción muscular en la parte del cuerpo representada. Este "somatotópico" Sin embargo, la representación no está distribuida uniformemente; la cabeza, por ejemplo, está representada por una región aproximadamente tres veces más grande que la zona de toda la espalda y el tronco. El tamaño de cualquier zona se correlaciona con la precisión del control motor y la discriminación sensorial posible. Las áreas de los labios, los dedos y la lengua son particularmente grandes, considerando el tamaño proporcional de las partes del cuerpo representadas.
Los mapas de áreas visuales son retinotópicos, lo que significa que reflejan la topografía de la retina: la capa de neuronas activadas por la luz que recubre la parte posterior del ojo. También en este caso la representación es desigual: la fóvea (la zona situada en el centro del campo visual) está muy sobrerrepresentada en comparación con la periferia. El circuito visual de la corteza cerebral humana contiene varias docenas de mapas retinotópicos distintos, cada uno de ellos dedicado a analizar el flujo de entrada visual de una manera particular. La corteza visual primaria (área 17 de Brodmann), que es el principal receptor de información directa desde la parte visual del tálamo, contiene muchas neuronas que se activan más fácilmente mediante bordes con una orientación particular que se mueven a través de un punto particular en el campo visual. Las áreas visuales situadas más abajo extraen características como el color, el movimiento y la forma.
En las áreas auditivas, el mapa primario es tonotópico. Los sonidos se analizan según la frecuencia (es decir, tono alto versus tono bajo) por áreas auditivas subcorticales, y este análisis se refleja en la zona auditiva primaria de la corteza. Al igual que ocurre con el sistema visual, existen varios mapas corticales tonotópicos, cada uno de ellos dedicado a analizar el sonido de una manera particular.
Dentro de un mapa topográfico a veces puede haber niveles más finos de estructura espacial. En la corteza visual primaria, por ejemplo, donde la organización principal es retinotópica y las principales respuestas son a los bordes en movimiento, las células que responden a diferentes orientaciones de los bordes están espacialmente segregadas unas de otras.
Para humanos y otros primates
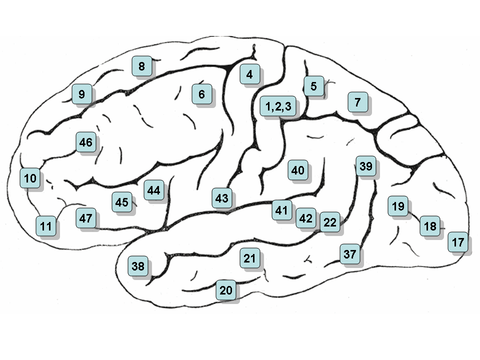
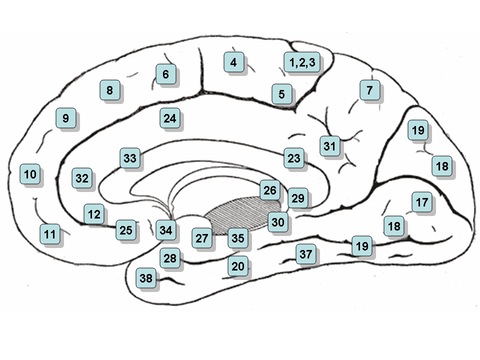
- Áreas 3, 1 y 2 - Corteza somatosensora primaria en el giro postcentral (frecuentemente denominado Áreas 3, 1, 2 por convención)
- Zona 4 – Corteza de motor primaria
- Zona 5 – Lobulo parietal superior
- Zona 6 – Corteza premotor y Cortex de motor suplementario (Secondary Motor Cortex) (zona de motor suplementaria)
- Zona 7 – Coordinación Visuo-Motor
- Zona 8 – Incluye campos de ojos frontales
- Zona 9 – Corteza prefrontal Dorsolateral
- Zona 10 – Corteza prefrontal anterior (la mayor parte rostral de giro frontal superior y medio)
- Zona 11 – Zona Orbitofrontal (orbital y recto gyri, además de parte de la parte rostral del giro frontal superior)
- Zona 12 – Zona Orbitofrontal (utilizada para formar parte de BA11, se refiere a la zona entre el giro frontal superior y el sulcus rostral inferior)
- Zona 13 y Zona 14* – Corteza insular
- Zona 15* – Lóbulo temporal anterior
- Zona 16 – Corteza insular
- Zona 17 – Corteza visual primaria (V1)
- Área 18 – Corteza visual secundaria (V2)
- Zona 19 – Corteza visual asociativa (V3, V4, V5)
- Zona 20 – Inferior giro temporal
- Zona 21 – Gris temporal medio
- Área 22 – Parte del giro temporal superior, incluido en el área de Wernicke
- Zona 23 – Corteza ventral posterior cingular
- Zona 24 – Corteza de cingulado anterior ventral.
- Zona 25 – Área subgénita (parte de la corteza prefrontal Ventromedial)
- Zona 26 – Porción eclesial de la región retrosplenial de la corteza cerebral
- Área 27 – Presubiculum
- Área 28 - Corteza ventral entorhinal
- Zona 29 – Corteza retrosplenial
- Zona 30 – Subdivisión de corteza retrosplenial
- Zona 31 - Dorsal Posterior cingulate cortex
- Área 32 - Dorsal anterior cingular cortex
- Área 33 – Parte de corteza cingular anterior
- Zona 34 – Dorsal entorhinal cortex (en el parahippocampal gyrus)
- Área 35 – Parte de la corteza perirhinal (en el sulcus rinoceronte)
- Área 36 – Parte de la corteza perirhinal (en el sulcus rinoceronte)
- Zona 37 - Fusiform gyrus
- Área 38 – Zona temporopolar (la parte más rostral del giro temporal superior y medio)
- Zona 39 – Gris angular, considerado por algunos como parte de la zona de Wernicke
- Zona 40 – Gris supramarginal considerado por algunos como parte de la zona de Wernicke
- Áreas 41 y 42 – Corteza Auditoria
- Área 43 - Corteza gustativa primaria
- Áreas 44 y 45 – El área de Broca, incluye la parte opercular y la parte triangular del giro frontal inferior
- Área 46 – Corteza prefrontal Dorsolateral
- Zona 47 – Parte orbital del giro frontal inferior
- Área 48 – Área retrosubicular (una pequeña parte de la superficie medial del lóbulo temporal)
- Área 49 – Área parasubicular en un roedor
- Área 52 – Área parainsular (en la unión del lóbulo temporal y la insula)
(*) Área que solo se encuentra en primates no humanos.
Algunas de las áreas originales de Brodmann se han subdividido aún más, por ejemplo, "23a" y "23b".
Mapa en el que se puede hacer clic: superficie lateral
- Nota: la vista lateral, o la vista lateral, del cerebro se denota la 'superficie lateral'
Mapa en el que se puede hacer clic: superficie media
- Nota: la vista de la sección entre los hemisferios derecho e izquierdo del cerebro se denota la "superficie media"
Crítica
Cuando von Bonin y Bailey construyeron un mapa cerebral para el mono macaco, encontraron inadecuada la descripción de Brodmann y escribieron: "Brodmann (1907), es cierto, preparó un mapa del cerebro humano que ha sido ampliamente reproducido, pero, lamentablemente, los datos en los que se basa nunca fueron publicados" En su lugar, utilizaron el esquema citoarquitectónico de Constantin von Economo y Georg N. Koskinas publicado en 1925, que tenía la "única descripción detallada aceptable de la corteza humana".
Contenido relacionado
Ley de Fick
Precisión y exactitud
Evidencia empírica