
Vuelo de insectos

Los insectos son el único grupo de invertebrados que han desarrollado alas y vuelo. Los insectos volaron por primera vez en el Carbonífero, hace entre 300 y 350 millones de años, lo que los convirtió en los primeros animales en desarrollar el vuelo. Las alas pueden haber evolucionado a partir de apéndices a los lados de las extremidades existentes, que ya tenían nervios, articulaciones y músculos utilizados para otros fines. Es posible que inicialmente se hayan utilizado para navegar sobre el agua o para reducir la velocidad de descenso al planear.
Dos grupos de insectos, las libélulas y las efímeras, tienen músculos de vuelo unidos directamente a las alas. En otros insectos alados, los músculos de vuelo se unen al tórax, lo que lo hace oscilar para inducir el batir de las alas. De estos insectos, algunos (moscas y algunos escarabajos) alcanzan frecuencias de aleteo muy altas mediante la evolución de un sistema "asincrónico" sistema nervioso, en el que el tórax oscila más rápido que la frecuencia de los impulsos nerviosos.
No todos los insectos son capaces de volar. Varios insectos ápteros han perdido secundariamente sus alas a través de la evolución, mientras que otros insectos más básicos, como los lepismas, nunca desarrollaron alas. En algunos insectos eusociales como las hormigas y las termitas, sólo las castas reproductivas tardías desarrollan alas durante la temporada de apareamiento antes de perderlas después del apareamiento, mientras que los miembros de otras castas no tienen alas durante toda su vida.
Algunos insectos muy pequeños no utilizan la aerodinámica de estado estacionario, sino el mecanismo de aplauso y lanzamiento de Weis-Fogh, generando grandes fuerzas de sustentación a expensas del desgaste de las alas. Muchos insectos pueden flotar, manteniendo la altura y controlando su posición. Algunos insectos, como las polillas, tienen las alas anteriores acopladas a las traseras para que puedan funcionar al unísono.
Mecanismos
Vuelo directo
A diferencia de otros insectos, los músculos de las alas de Ephemeroptera (efímeras) y Odonata (libélulas y caballitos del diablo) se insertan directamente en las bases de las alas, que están articuladas de modo que un pequeño movimiento hacia abajo de la base del ala eleva el ala hacia arriba, mucho como remar por el aire. Las libélulas y los caballitos del diablo tienen alas delanteras y traseras similares en forma y tamaño. Cada uno opera de forma independiente, lo que les da un grado de control y movilidad finos en términos de la brusquedad con la que pueden cambiar de dirección y velocidad, algo que no se ve en otros insectos voladores. Los odonatos son todos depredadores aéreos y siempre han cazado otros insectos en el aire.
 Vuelo directo: músculos unidos a alas. Insectos grandes sólo
Vuelo directo: músculos unidos a alas. Insectos grandes sólo Los Odonata (dragonflies y damselflies) tienen musculatura de vuelo directa, como los mayflies.
Los Odonata (dragonflies y damselflies) tienen musculatura de vuelo directa, como los mayflies.- Vuelo de movimiento lento de libélula


Vuelo indirecto
Aparte de los dos órdenes con músculos de vuelo directo, todos los demás insectos alados vivos vuelan utilizando un mecanismo diferente, que involucra músculos de vuelo indirecto. Este mecanismo evolucionó una vez y es la característica definitoria (sinapomorfía) de la infraclase Neoptera; Se corresponde, probablemente no por coincidencia, con la aparición de un mecanismo de plegado de las alas, que permite a los insectos neopteranos plegar las alas hacia atrás sobre el abdomen cuando están en reposo (aunque esta capacidad se ha perdido de forma secundaria en algunos grupos, como en las mariposas). .
Sin embargo, lo que todos los Neoptera comparten es la forma en que funcionan los músculos del tórax: estos músculos, en lugar de unirse a las alas, se adhieren al tórax y lo deforman; Dado que las alas son extensiones del exoesqueleto torácico, las deformaciones del tórax hacen que las alas también se muevan. Un conjunto de músculos longitudinales a lo largo de la espalda comprime el tórax de adelante hacia atrás, lo que hace que la superficie dorsal del tórax (notum) se doble hacia arriba, lo que hace que las alas giren hacia abajo. Otro conjunto de músculos desde el tergum hasta el esternón tira del notum hacia abajo nuevamente, lo que hace que las alas se giren hacia arriba.
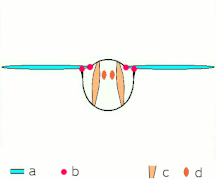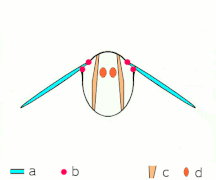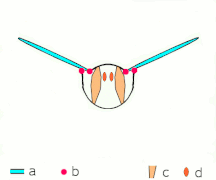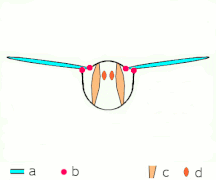 Vuelo indirecto: los músculos hacen que el tórax oscila en la mayoría de los insectos
Vuelo indirecto: los músculos hacen que el tórax oscila en la mayoría de los insectos El Neoptera, incluyendo mariposas y la mayoría de insectos, tienen musculatura de vuelo indirecta
El Neoptera, incluyendo mariposas y la mayoría de insectos, tienen musculatura de vuelo indirecta


Los insectos que baten sus alas menos de cien veces por segundo utilizan músculos sincrónicos. El músculo sincrónico es un tipo de músculo que se contrae una vez por cada impulso nervioso. Esto generalmente produce menos potencia y es menos eficiente que el músculo asincrónico, lo que explica la evolución independiente de los músculos de vuelo asincrónicos en varios clados de insectos separados.
Los insectos que baten sus alas más rápidamente, como el abejorro, utilizan músculos asincrónicos; este es un tipo de músculo que se contrae más de una vez por impulso nervioso. Esto se logra estimulando el músculo para que se contraiga nuevamente mediante una liberación de tensión en el músculo, lo que puede ocurrir más rápidamente que mediante una simple estimulación nerviosa. Esto permite que la frecuencia de los batidos de las alas supere la velocidad a la que el sistema nervioso puede enviar impulsos. El músculo asincrónico es uno de los últimos refinamientos que ha aparecido en algunos de los neópteros superiores (coleópteros, dípteros e himenópteros). El efecto general es que muchos Neoptera superiores pueden batir sus alas mucho más rápido que los insectos con músculos de vuelo directo.
El músculo asincrónico está, por definición, bajo un control relativamente aproximado por parte del sistema nervioso. Para equilibrar esta compensación evolutiva, los insectos que evolucionaron en vuelo indirecto también desarrollaron un sistema neuromuscular separado para el control detallado del aleteo. Conocidos como "músculos directos", estos músculos se unen directamente a los escleritos que forman la bisagra del ala y se contraen con impulsos 1:1 de las neuronas motoras. Trabajos recientes han comenzado a abordar la compleja dinámica muscular no lineal en la bisagra del ala y sus efectos en la trayectoria de la punta del ala.
Aerodinámica
Hay dos modelos aerodinámicos básicos del vuelo de los insectos: crear un vórtice de vanguardia y utilizar aplaudir y lanzar.
Vórtice de vanguardia
La mayoría de los insectos utilizan un método que crea un vórtice de borde de ataque en espiral. Estas alas batientes se mueven mediante dos medios movimientos básicos. El movimiento descendente comienza hacia arriba y hacia atrás y se hunde hacia abajo y hacia adelante. Luego, el ala se gira rápidamente (supinación) de modo que el borde de ataque apunte hacia atrás. Luego, la carrera ascendente empuja el ala hacia arriba y hacia atrás. Luego, el ala se gira nuevamente (pronación) y puede ocurrir otro movimiento descendente. El rango de frecuencia en insectos con músculos de vuelo sincrónicos suele ser de 5 a 200 hercios (Hz). En aquellos con músculos de vuelo asincrónicos, la frecuencia de aleteo puede exceder los 1000 Hz. Cuando el insecto está flotando, los dos movimientos duran el mismo tiempo. Sin embargo, un movimiento descendente más lento proporciona empuje.
La identificación de las fuerzas principales es fundamental para comprender el vuelo de los insectos. Los primeros intentos de comprender el aleteo asumieron un estado casi estacionario. Esto significa que se supuso que el flujo de aire sobre el ala en un momento dado era el mismo que sería sobre un ala en estado estacionario y sin aleteo con el mismo ángulo de ataque. Al dividir el ala batiente en una gran cantidad de posiciones inmóviles y luego analizar cada posición, sería posible crear una línea de tiempo de las fuerzas instantáneas sobre el ala en cada momento. Se descubrió que la sustentación calculada era demasiado pequeña en un factor de tres, por lo que los investigadores se dieron cuenta de que debía haber fenómenos inestables que proporcionaban fuerzas aerodinámicas. Se estaban desarrollando varios modelos analíticos que intentaban aproximar el flujo cerca del ala batiente. Algunos investigadores predijeron picos de fuerza en la supinación. Estas fuerzas predichas se confirmaron posteriormente con un modelo de mosca de la fruta a escala dinámica. Otros argumentaron que los picos de fuerza durante la supinación y la pronación son causados por un efecto rotacional desconocido que es fundamentalmente diferente de los fenómenos traslacionales. Hay cierto desacuerdo con este argumento. Mediante la dinámica de fluidos computacional, algunos investigadores sostienen que no existe un efecto de rotación. Afirman que las fuerzas elevadas se deben a una interacción con la estela dejada por el golpe anterior.
Al igual que el efecto de rotación mencionado anteriormente, los fenómenos asociados con el aleteo de las alas no se comprenden ni se acuerdan completamente. Debido a que cada modelo es una aproximación, diferentes modelos omiten efectos que se presumen insignificantes. Por ejemplo, el efecto Wagner, propuesto por Herbert A. Wagner en 1925, dice que la circulación aumenta lentamente hasta su estado estacionario debido a la viscosidad cuando un ala inclinada se acelera desde el reposo. Este fenómeno explicaría un valor de elevación inferior al previsto. Normalmente, el caso ha sido encontrar fuentes para el impulso adicional. Se ha argumentado que este efecto es insignificante para el flujo con un número de Reynolds típico del vuelo de los insectos. El número de Reynolds es una medida de turbulencia; El flujo es laminar (suave) cuando el número de Reynolds es bajo y turbulento cuando es alto. El efecto Wagner fue ignorado, conscientemente, al menos en un modelo. Uno de los fenómenos más importantes que se produce durante el vuelo de los insectos es la succión del borde de ataque. Esta fuerza es importante para el cálculo de la eficiencia. El concepto de succión de borde de ataque fue propuesto por primera vez por D. G. Ellis y J. L. Stollery en 1988 para describir la elevación de vórtice en alas delta de bordes afilados. En ángulos de ataque elevados, el flujo se separa sobre el borde de ataque, pero se vuelve a unir antes de llegar al borde de salida. Dentro de esta burbuja de flujo separado hay un vórtice. Debido a que el ángulo de ataque es tan alto, se transfiere una gran cantidad de impulso hacia abajo al flujo. Estas dos características crean una gran cantidad de fuerza de elevación, así como algo de resistencia adicional. Lo importante, sin embargo, es el ascensor. Debido a que el flujo se ha separado, pero todavía proporciona una gran cantidad de sustentación, este fenómeno se llama retardo de pérdida, observado por primera vez en las hélices de los aviones por H. Himmelskamp en 1945. Este efecto se observó en el vuelo de los insectos aleteando y se demostró que era capaz de proporcionando suficiente impulso para explicar la deficiencia en los modelos de estado cuasi estacionario. Este efecto lo utilizan los piragüistas en un golpe de remo.
Todos los efectos sobre el aleteo de un ala pueden reducirse a tres fuentes principales de fenómenos aerodinámicos: el vórtice del borde de ataque, las fuerzas aerodinámicas en estado estacionario sobre el ala y el contacto del ala con su estela de anteriores trazos. El tamaño de los insectos voladores oscila entre unos 20 microgramos y unos 3 gramos. A medida que aumenta la masa corporal del insecto, aumenta el área del ala y disminuye la frecuencia del aleteo. Para insectos más grandes, el número de Reynolds (Re) puede llegar a 10000, cuando el flujo comienza a volverse turbulento. Para los insectos más pequeños, puede ser tan bajo como 10. Esto significa que los efectos viscosos son mucho más importantes para los insectos más pequeños.


Otra característica interesante del vuelo de los insectos es la inclinación del cuerpo. A medida que aumenta la velocidad de vuelo, el cuerpo del insecto tiende a inclinarse hacia abajo y volverse más horizontal. Esto reduce el área frontal y por tanto, la resistencia del cuerpo. Dado que la resistencia también aumenta a medida que aumenta la velocidad de avance, el insecto hace que su vuelo sea más eficiente a medida que esta eficiencia se vuelve más necesaria. Además, al cambiar el ángulo geométrico de ataque en el movimiento descendente, el insecto puede mantener su vuelo con una eficiencia óptima mediante tantas maniobras como sea posible. El desarrollo del empuje general es relativamente pequeño en comparación con las fuerzas de sustentación. Las fuerzas de elevación pueden ser más de tres veces el peso del insecto, mientras que el empuje incluso a las velocidades más altas puede ser tan solo el 20% del peso. Esta fuerza se desarrolla principalmente a través del movimiento ascendente menos potente del movimiento de aleteo.
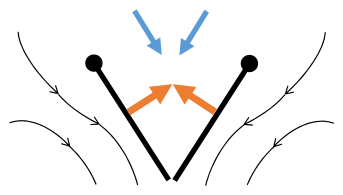
Aplaude y lanza


Aplaudir y lanzar, o el mecanismo de Weis-Fogh, descubierto por el zoólogo danés Torkel Weis-Fogh, es un método de generación de sustentación utilizado durante el vuelo de pequeños insectos. A medida que los tamaños de los insectos llegan a ser inferiores a 1 mm, las fuerzas viscosas se vuelven dominantes y la eficacia de la generación de sustentación desde un perfil aerodinámico disminuye drásticamente. A partir de la posición de palmada, las dos alas se separan y giran alrededor del borde de salida. Luego, las alas se separan y se desplazan horizontalmente hasta el final del recorrido descendente. A continuación, las alas se pronan y utilizan el borde de ataque durante un movimiento de remo ascendente. Cuando comienza el movimiento de aplauso, los bordes delanteros se encuentran y giran juntos hasta que el espacio desaparece. Inicialmente, se pensó que las alas se estaban tocando, pero varios incidentes indican un espacio entre las alas y sugieren que proporciona un beneficio aerodinámico.
La generación de elevación a partir del mecanismo de aplauso y lanzamiento ocurre durante varios procesos a lo largo del movimiento. En primer lugar, el mecanismo se basa en una interacción ala-ala, ya que un solo movimiento de ala no produce suficiente sustentación. A medida que las alas giran alrededor del borde de salida en el movimiento de lanzamiento, el aire se precipita hacia el espacio creado y genera un fuerte vórtice en el borde de ataque, y un segundo que se desarrolla en las puntas de las alas. Un tercer vórtice, más débil, se desarrolla en el borde de salida. La fuerza de los vórtices en desarrollo depende, en parte, de la brecha inicial de la separación entre las alas al inicio del movimiento de lanzamiento. Con una brecha reducida entre las alas, lo que indica una mayor generación de sustentación, a costa de mayores fuerzas de arrastre. Se descubrió que la implementación de un movimiento de agitación durante el lanzamiento, alas flexibles y un mecanismo de pérdida retardada refuerzan la estabilidad y la fijación del vórtice. Finalmente, para compensar la menor producción general de sustentación durante el vuelo con un número de Reynolds bajo (con flujo laminar), los insectos pequeños a menudo tienen una frecuencia de carrera más alta para generar velocidades en las puntas de las alas que son comparables a las de los insectos más grandes.
Las fuerzas de arrastre más grandes esperadas ocurren durante el movimiento de lanzamiento dorsal, ya que las alas necesitan separarse y girar. La atenuación de las grandes fuerzas de arrastre se produce mediante varios mecanismos. Se descubrió que las alas flexibles disminuyen la resistencia en el movimiento de lanzamiento hasta en un 50% y reducen aún más la resistencia general durante toda la carrera del ala en comparación con las alas rígidas. Las cerdas en los bordes de las alas, como se ve en Encarsia formosa, provocan una porosidad en el flujo que aumenta y reduce las fuerzas de arrastre, a costa de una menor generación de sustentación. Además, la separación entre alas antes del lanzamiento juega un papel importante en el efecto general de la resistencia. A medida que aumenta la distancia entre las alas, la resistencia total disminuye.
El mecanismo de aplaudir y lanzar también lo utiliza el molusco marino Limacina helicina, una mariposa marina. Algunos insectos, como el minador vegetal Liriomyza sativae (una mosca), aprovechan un aplauso y un lanzamiento parciales, utilizando el mecanismo solo en la parte exterior del ala para aumentar la sustentación en aproximadamente un 7% cuando flotan. .
- Mecanismo de vuelo de solapa y vuelo después de Sane 2003
 Clap 1: alas cerca de atrás
Clap 1: alas cerca de atrás Clap 2: toque de los bordes principales, el ala gira alrededor del borde principal, forma de vórtices
Clap 2: toque de los bordes principales, el ala gira alrededor del borde principal, forma de vórtices Clap 3: bordes de seguimiento cerca, vórtices cobertizo, alas cerca dando empuje
Clap 3: bordes de seguimiento cerca, vórtices cobertizo, alas cerca dando empuje



- Flechas negras: flujo; flechas azules: velocidad inducida; Flechas naranjas: fuerza neta en el ala
 Fling 1: las alas giran alrededor del borde del sendero para aflojar
Fling 1: las alas giran alrededor del borde del sendero para aflojar Fling 2: el borde líder se aleja, el aire se precipita, aumento del ascensor
Fling 2: el borde líder se aleja, el aire se precipita, aumento del ascensor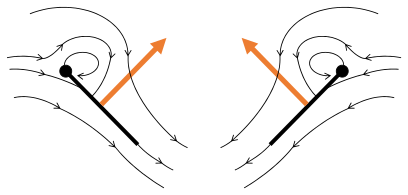 Fling 3: nuevas formas de vórtice a la vanguardia, los vórtices de bordes que siguen se anulan, tal vez ayudando a que el flujo crezca más rápido (Weis-Fogh 1973)
Fling 3: nuevas formas de vórtice a la vanguardia, los vórtices de bordes que siguen se anulan, tal vez ayudando a que el flujo crezca más rápido (Weis-Fogh 1973)



Ecuaciones rectoras
Un ala que se mueve en fluidos experimenta una fuerza fluida, que sigue las convenciones que se encuentran en la aerodinámica. La componente de fuerza normal a la dirección del flujo relativa al ala se llama sustentación (L), y la componente de fuerza en la dirección opuesta del flujo es resistencia (D ). Con los números de Reynolds considerados aquí, una unidad de fuerza apropiada es 1/2(ρU2S), donde ρ es la densidad del fluido, S el área del ala y U la velocidad del ala. Las fuerzas adimensionales se denominan coeficientes de sustentación (CL) y de arrastre (CD), es decir:

CL y CD son constantes sólo si el flujo es estable. Una clase especial de objetos, como los perfiles aerodinámicos, pueden alcanzar un estado estable cuando cortan el fluido con un pequeño ángulo de ataque. En este caso, el flujo no viscoso alrededor de un perfil aerodinámico puede aproximarse mediante un flujo potencial que satisface la condición límite de no penetración. El teorema de Kutta-Joukowski de un perfil aerodinámico 2D supone además que el flujo sale suavemente del borde de salida afilado, y esto determina la circulación total alrededor de un perfil aerodinámico. La elevación correspondiente viene dada por el principio de Bernoulli (teorema de Blasius):

Los flujos alrededor de aves e insectos pueden considerarse incompresibles: el número de Mach, o velocidad relativa a la velocidad del sonido en el aire, suele ser 1/300 y la frecuencia del ala es de aproximadamente 10 a 103 Hz. Usando la ecuación gobernante como la ecuación de Navier-Stokes sujeta a la condición de frontera de no deslizamiento, la ecuación es:

Donde u(x, t) es el campo de flujo, p la presión, ρ la densidad del fluido, ν la viscosidad cinemática, ubd la velocidad a el límite y us la velocidad del sólido. Al elegir una escala de longitud, L, y una escala de velocidad, U, la ecuación se puede expresar en forma adimensional que contiene el número de Reynolds, Re=uL/ν. Hay dos diferencias obvias entre el ala de un insecto y un perfil aerodinámico: el ala de un insecto es mucho más pequeña y aletea. Usando una libélula como ejemplo, su cuerda (c) es de aproximadamente 1 cm (0,39 pulgadas), la longitud de su ala (l) de aproximadamente 4 cm (1,6 pulgadas) y su frecuencia de ala (f) de aproximadamente 40 Hz. La velocidad punta (u) es de aproximadamente 1 m/s (3,3 pies/s) y el número de Reynolds correspondiente es de aproximadamente 103. En el extremo más pequeño, una avispa calcidoide típica tiene una longitud de ala de aproximadamente 0,5 a 0,7 mm (0,020 a 0,028 mm). in) y bate sus alas a unos 400 Hz. Su número de Reynolds es aproximadamente 25. El rango del número de Reynolds en el vuelo de los insectos es de aproximadamente 10 a 104, que se encuentra entre los dos límites que son convenientes para las teorías: flujos constantes invisibles alrededor de un perfil aerodinámico y Stokes. flujo experimentado por una bacteria nadadora. Por este motivo, este rango intermedio no se comprende bien. Por otro lado, es quizás el régimen más omnipresente entre las cosas que vemos. Las hojas y semillas que caen, los peces y los pájaros encuentran flujos inestables similares a los que se ven alrededor de un insecto. El número de Reynolds en sentido de cuerda se puede describir mediante:

y


Donde es la longitud media del acorde, es la velocidad de la punta del ala, es la amplitud del golpe, es la frecuencia de los golpes, es el radio de la hidratación, es el área del ala, y es la longitud del ala, incluyendo la punta del ala.







Además del número de Reynolds, existen al menos otros dos parámetros adimensionales relevantes. Un ala tiene tres escalas de velocidad: la velocidad de aleteo con respecto al cuerpo (u), la velocidad de avance del cuerpo (U0) y la velocidad de lanzamiento (Ωc). Las proporciones de ellos forman dos variables adimensionales, U0/u y Ωc/u; la primera suele ser Se conoce como relación de avance y también está relacionada con la frecuencia reducida, fc/U0.
Si el ala de un insecto es rígida, por ejemplo, el ala de una Drosophila lo es aproximadamente, su movimiento relativo a un cuerpo fijo puede describirse mediante tres variables: la posición de la punta en coordenadas esféricas, (Θ(t),Φ(t)), y el ángulo de cabeceo ψ(t), alrededor del eje que conecta la raíz y la punta. Para estimar las fuerzas aerodinámicas basándose en el análisis de elementos de pala, también es necesario determinar el ángulo de ataque (α). El ángulo de ataque típico con una envergadura del 70% varía de 25° a 45° en insectos flotantes (15° en colibríes). A pesar de la gran cantidad de datos disponibles para muchos insectos, relativamente pocos experimentos informan la variación temporal de α durante un golpe. Entre ellos se encuentran experimentos en túneles de viento con una langosta atada y una mosca atada, y el vuelo libre de una mosca de la fruta.
Debido a que son relativamente fáciles de medir, las trayectorias de las puntas de las alas se han informado con mayor frecuencia. Por ejemplo, seleccionar solo secuencias de vuelo que produjeron suficiente sustentación para soportar un peso mostrará que la punta del ala sigue una forma elíptica. También se informaron formas no cruzadas para otros insectos. Independientemente de sus formas exactas, el movimiento de obstrucción indica que los insectos pueden usar la resistencia aerodinámica además de la elevación para soportar su peso.
Flotando
| Parámetros de vuelo | Velocidad (m/s) | Beats/s |
|---|---|---|
| Aeshnid Dragonfly | 7.0 | 38 |
| Hornet | 5.7 | 100 |
| Hummingbird hawkmoth | 5.0 | 85 |
| caballo | 3.9 | 96 |
| Hoverfly | 3.5 | 120 |
| Bumblebee | 2.9 | 130 |
| Honeybee | 2.5 | 250 |
| Housefly | 2.0 | 190 |
| Presuntamente | 1,5 | 16 |
| Escorpión | 0.49 | 28 |
| mariposa blanca grande | 2.5 | 12 |
| Umbral (clap y fling) | 0.3 | 254 |
.jpg)
Muchos insectos pueden flotar o permanecer en un lugar en el aire, batiendo sus alas rápidamente. Hacerlo requiere estabilización lateral así como producción de sustentación. La fuerza de elevación se produce principalmente durante la carrera descendente. A medida que las alas empujan hacia abajo el aire circundante, la fuerza de reacción resultante del aire sobre las alas empuja al insecto hacia arriba. Las alas de la mayoría de los insectos están evolucionadas de modo que, durante el movimiento hacia arriba, la fuerza sobre el ala es pequeña. Dado que el golpe fuerte y el golpe de retorno fuerzan al insecto hacia arriba y hacia abajo respectivamente, el insecto oscila y termina permaneciendo en la misma posición.
La distancia que cae el insecto entre aleteos depende de qué tan rápido baten sus alas: cuanto más lento aletea, más largo es el intervalo en el que cae y más lejos cae entre cada aleteo. Se puede calcular la frecuencia de aleteo necesaria para que el insecto mantenga una determinada estabilidad en su amplitud. Para simplificar los cálculos, se debe suponer que la fuerza de elevación tiene un valor constante finito mientras las alas se mueven hacia abajo y que es cero mientras las alas se mueven hacia arriba. Durante el intervalo de tiempo Δt del aleteo ascendente, el insecto cae una distancia h bajo la influencia de la gravedad.

El trazo hacia arriba restaura el insecto a su posición original. Normalmente, puede ser necesario que la posición vertical del insecto cambie no más de 0,1 mm (es decir, h = 0,1 mm). El tiempo máximo permitido para la caída libre es entonces

Dado que los movimientos hacia arriba y hacia abajo de las alas tienen aproximadamente la misma duración, el período T para un ala completa hacia arriba y hacia abajo es dos veces Δr, es decir,

La frecuencia de los latidos, f, es decir, el número de aleteos por segundo, está representada por la ecuación:

En los ejemplos utilizados, la frecuencia utilizada es 110 latidos/s, que es la frecuencia típica que se encuentra en los insectos. Las mariposas tienen una frecuencia mucho más lenta, alrededor de 10 latidos/s, lo que significa que no pueden flotar. Otros insectos pueden producir una frecuencia de 1000 latidos/s. Para restaurar el insecto a su posición vertical original, la fuerza promedio hacia arriba durante el movimiento hacia abajo, Fav, debe ser igual al doble del peso del insecto. Tenga en cuenta que, dado que la fuerza ascendente sobre el cuerpo del insecto se aplica sólo durante la mitad del tiempo, la fuerza ascendente promedio sobre el insecto es simplemente su peso.
Entrada de energía
Ahora se puede calcular la potencia necesaria para mantenerse flotando, considerando nuevamente un insecto con masa m 0,1 g, la fuerza promedio, Fav, aplicada por las dos alas durante el movimiento hacia abajo es dos veces el peso. Debido a que la presión aplicada por las alas se distribuye uniformemente sobre el área total del ala, eso significa que se puede suponer que la fuerza generada por cada ala actúa a través de un solo punto en la sección media de las alas. Durante el movimiento descendente, el centro de las alas recorre una distancia vertical d. El trabajo total realizado por el insecto durante cada golpe hacia abajo es el producto de la fuerza y la distancia; eso es,

Si las alas se mueven siguiendo el ritmo en un ángulo de 70°, entonces, en el caso presentado para el insecto con alas de 1 cm de largo, d es 0,57 cm. Por tanto, el trabajo realizado durante cada golpe por las dos alas es:

La energía se utiliza para elevar al insecto contra la gravedad. La energía E necesaria para elevar la masa del insecto 0,1 mm durante cada movimiento descendente es:

Esta es una fracción insignificante de la energía total gastada y, claramente, la mayor parte de la energía se gasta en otros procesos. Un análisis más detallado del problema muestra que el trabajo realizado por las alas se convierte principalmente en energía cinética del aire que se acelera con el movimiento descendente de las alas. La potencia es la cantidad de trabajo realizado en 1 s; en el ejemplo del insecto, realiza 110 movimientos hacia abajo por segundo. Por lo tanto, su potencia de salida P es, golpes por segundo, y eso significa que su potencia de salida P es:

Potencia de salida
En el cálculo de la potencia utilizada en el vuelo estacionario, los ejemplos utilizados ignoraron la energía cinética de las alas en movimiento. Las alas de los insectos, por ligeras que sean, tienen una masa finita; por lo tanto, a medida que se mueven poseen energía cinética. Debido a que las alas están en movimiento giratorio, la energía cinética máxima durante cada golpe del ala es:

Aquí I es el momento de inercia del ala y ωmax es la velocidad angular máxima durante el recorrido del ala. Para obtener el momento de inercia del ala, supondremos que el ala se puede aproximar mediante una varilla delgada que gira en un extremo. Entonces el momento de inercia del ala es:

Donde l es la longitud del ala (1 cm) y m es la masa de dos alas, que normalmente puede ser 10−3 g. La velocidad angular máxima, ωmax, se puede calcular a partir de la velocidad lineal máxima, νmax, en el centro del ala:

Durante cada golpe, el centro de las alas se mueve con una velocidad lineal promedio νav dada por la distancia d recorrida por el centro de el ala dividida por la duración Δt de la carrera del ala. De nuestro ejemplo anterior, d = 0,57 cm y Δt = 4,5×10−3 s. Por lo tanto:

La velocidad de las alas es cero tanto al principio como al final del recorrido del ala, lo que significa que la velocidad lineal máxima es mayor que la velocidad promedio. Si suponemos que la velocidad oscila (sinusoidalmente) a lo largo de la trayectoria del ala, la velocidad máxima es el doble que la velocidad promedio. Por tanto, la velocidad angular máxima es:

Y la energía cinética por tanto es:

Dado que hay dos carreras de ala (la carrera ascendente y la carrera descendente) en cada ciclo del movimiento del ala, la energía cinética es 2×43 = 86 erg. Esto es aproximadamente tanta energía como la que se consume al flotar.
Elasticidad

Los insectos obtienen energía cinética, proporcionada por los músculos, cuando las alas aceleran. Cuando las alas comienzan a desacelerar hacia el final del recorrido, esta energía debe disiparse. Durante la brazada, la energía cinética es disipada por los propios músculos y se convierte en calor (este calor a veces se utiliza para mantener la temperatura corporal central). Algunos insectos pueden utilizar la energía cinética del movimiento ascendente de las alas para ayudar en su vuelo. Las articulaciones de las alas de estos insectos contienen una almohadilla de proteína elástica parecida al caucho llamada resilina. Durante el ascenso del ala, la resilina se estira. La energía cinética del ala se convierte en energía potencial en la resilina estirada, que almacena la energía de forma muy parecida a un resorte. Cuando el ala desciende, esta energía se libera y ayuda en el descenso.
Utilizando algunas suposiciones simplificadoras, podemos calcular la cantidad de energía almacenada en la resilina estirada. Aunque la resilina está doblada para darle una forma compleja, el ejemplo dado muestra el cálculo como una varilla recta de área A y longitud. Además, asumiremos que durante todo el tramo la resilina obedece la ley de Hooke. Esto no es estrictamente cierto ya que la resilina se estira en una cantidad considerable y, por lo tanto, tanto el área como el módulo de Young cambian en el proceso de estiramiento. La energía potencial U almacenada en la resilina estirada es:

Aquí E es el módulo de Young para la resilina, que se ha medido en 1,8×107 dyn/cm2. Normalmente, en un insecto del tamaño de una abeja, el volumen de resilina puede ser equivalente a un cilindro de 2×10−2 cm de largo y 4×10−4 cm< sup>2 en área. En el ejemplo dado, la longitud de la varilla de resilina aumenta en un 50% cuando se estira. Es decir, Δℓ es 10−2 cm. Por tanto, en este caso la energía potencial almacenada en la resilina de cada ala es:

La energía almacenada en las dos alas de un insecto del tamaño de una abeja es de 36 ergios, que es comparable a la energía cinética en el movimiento ascendente de las alas. Los experimentos muestran que hasta el 80% de la energía cinética del ala puede almacenarse en la resilina.
Acoplamiento de ala

Algunos órdenes de insectos de cuatro alas, como los lepidópteros, han desarrollado mecanismos morfológicos de acoplamiento de alas en la imago que hacen que estos taxones sean funcionalmente de dos alas. Todas las formas, excepto las más basales, exhiben este acoplamiento de alas.
Los mecanismos son de tres tipos diferentes: yugal, frenulo-retinacular y amplexiforme:
- Los grupos más primitivos tienen un área agrandada como el lóbulo cerca del margen basal posterior, es decir, en la base de la horca, un jugum, que se dobla bajo la caminata en vuelo.
- Otros grupos tienen un frenulum en la hindwing que se engancha bajo un retinaculo en el faldeo.
- En casi todas las mariposas y en la Bombycoidea (excepto la Sphingidae), no hay arreglo de frenulum y retinaculum para acoplar las alas. En su lugar, un área humeral ampliada de la hindú está ampliamente superpuesta por la proa. A pesar de la ausencia de una conexión mecánica específica, las alas se superponen y operan en fase. El derrame de poder del desfiladero empuja hacia abajo la marcha al unísono. Este tipo de acoplamiento es una variación de tipo frenado pero donde el frenulum y el retinaculo se pierden completamente.
Bioquímica
La bioquímica del vuelo de los insectos ha sido objeto de considerables estudios. Mientras que muchos insectos utilizan carbohidratos y lípidos como fuente de energía para volar, muchos escarabajos y moscas utilizan el aminoácido prolina como fuente de energía. Algunas especies también utilizan una combinación de fuentes y polillas como Manduca sexta utilizan carbohidratos para el calentamiento previo al vuelo.
Retroalimentación sensorial
Los insectos utilizan la retroalimentación sensorial para mantener y controlar el vuelo. Las investigaciones han demostrado el papel de estructuras sensoriales como antenas, halterios y alas en el control de la postura de vuelo, la amplitud y la frecuencia de los batidos.
Evolución y adaptación

En algún momento del período Carbonífero, hace entre 350 y 300 millones de años, cuando sólo había dos masas terrestres importantes, los insectos comenzaron a volar. Entre los fósiles de insectos alados más antiguos se encuentra Delitzschala, un paleodictióptero del Carbonífero Inferior; Rhyniognatha es más antiguo, del Devónico temprano, pero no se sabe si tenía alas o si era un insecto.
No se comprende bien cómo y por qué se desarrollaron las alas de los insectos, en gran parte debido a la escasez de fósiles apropiados del período de su desarrollo en el Carbonífero Inferior. Históricamente ha habido tres teorías principales sobre los orígenes del vuelo de los insectos. La primera fue que son modificaciones de branquias abdominales móviles, como las que se encuentran en las náyades acuáticas de las moscas efímeras. El análisis filogenómico sugiere que los Polyneoptera, el grupo de insectos alados que incluye a los saltamontes, evolucionaron a partir de un ancestro terrestre, lo que hace poco probable la evolución de las alas a partir de las branquias. Un estudio adicional sobre el comportamiento de salto de las larvas de efímera ha determinado que las branquias traqueales no desempeñan ningún papel en la guía del descenso de los insectos, lo que proporciona más evidencia en contra de esta hipótesis evolutiva. Esto deja dos teorías históricas importantes: que las alas se desarrollaron a partir de lóbulos paranotales, extensiones de los tergos torácicos; o que surgieron de modificaciones de segmentos de las piernas, que ya contenían músculos.
Hipótesis epicoxal (branquias abdominales)
.JPG)
Numerosos entomólogos, incluidos Landois en 1871, Lubbock en 1873, Graber en 1877 y Osborn en 1905, han sugerido que un posible origen de las alas de los insectos podrían haber sido las branquias abdominales móviles que se encuentran en muchos insectos acuáticos, como las náyades de las moscas efímeras. Según esta teoría estas branquias traqueales, que comenzaron su camino como salidas del sistema respiratorio y con el tiempo fueron modificadas con fines locomotores, con el tiempo se convirtieron en alas. Las branquias traqueales están equipadas con pequeñas aletas que vibran perpetuamente y tienen sus propios pequeños músculos rectos.
Hipótesis paranotal (tergal)
La hipótesis del lóbulo paranotal o tergal (pared dorsal del cuerpo), propuesta por Fritz Müller en 1875 y reelaborada por G. Crampton en 1916, Jarmila Kukalova-Peck en 1978 y Alexander P. Rasnitsyn en 1981, entre otros, sugiere que el insecto& Las alas de #39;se desarrollaron a partir de lóbulos paranotales, una preadaptación encontrada en fósiles de insectos que habría ayudado a la estabilización al saltar o caer. A favor de esta hipótesis está la tendencia de la mayoría de los insectos, cuando se asustan mientras trepan por las ramas, a escapar dejándose caer al suelo. Estos lóbulos habrían servido como paracaídas y permitirían al insecto aterrizar más suavemente. La teoría sugiere que estos lóbulos crecieron gradualmente y en una etapa posterior desarrollaron una articulación con el tórax. Incluso más tarde aparecerían los músculos para mover estas toscas alas. Este modelo implica un aumento progresivo de la efectividad de las alas, comenzando con el paracaidismo, luego el planeo y finalmente el vuelo activo. Aún así, la falta de evidencia fósil sustancial del desarrollo de las articulaciones y los músculos de las alas plantea una dificultad importante para la teoría, al igual que el desarrollo aparentemente espontáneo de la articulación y la venación, y ha sido rechazada en gran medida por los expertos en el campo.
Hipótesis endita-éxita (pleural)

En 1990, J. W. H. Trueman propuso que el ala se adaptó a partir de endites y exites, apéndices en las respectivas caras interna y externa de la extremidad del artrópodo primitivo, también llamada hipótesis pleural. Esto se basó en un estudio realizado por Goldschmidt en 1945 sobre Drosophila melanogaster, en el que una variación llamada "pod" (para podómeros, segmentos de extremidades) mostraron una mutación que transformó las alas normales. El resultado se interpretó como una disposición de patas de triple articulación con algunos apéndices adicionales pero sin el tarso, donde normalmente estaría la superficie costal del ala. Esta mutación fue reinterpretada como una fuerte evidencia de una fusión de salida dorsal y endita, en lugar de una pierna, y los apéndices encajan mucho mejor con esta hipótesis. La inervación, articulación y musculatura necesarias para la evolución de las alas ya están presentes en los segmentos de las extremidades.
Otras hipótesis
Otras hipótesis incluyen la sugerencia de Vincent Wigglesworth de 1973 de que las alas se desarrollaron a partir de protuberancias torácicas utilizadas como radiadores.
Adrian Thomas y Åke Norberg sugirieron en 2003 que las alas pueden haber evolucionado inicialmente para navegar en la superficie del agua, como se ve en algunas moscas de piedra.
Stephen P. Yanoviak y sus colegas propusieron en 2009 que el ala se deriva de un descenso aéreo dirigido, un fenómeno previo al vuelo que se encuentra en algunos apterygota, un taxón hermano sin alas de los insectos alados.
Doble origen
Biólogos como Averof, Niwa, Elias-Neto y sus colegas han comenzado a explorar el origen del ala del insecto utilizando evo-devo además de evidencia paleontológica. Esto sugiere que las alas son serialmente homólogas con las estructuras tergal y pleural, lo que potencialmente resuelve el debate de siglos de antigüedad. Jakub Prokop y sus colegas encontraron en 2017 evidencia paleontológica en las almohadillas de las alas de ninfas del Paleozoico de que las alas tenían un origen tan dual.