
Vida marina


Vida marina, vida marina o vida oceánica son las plantas, animales y otros organismos que viven en el agua salada de los mares. u océanos, o el agua salobre de los estuarios costeros. A un nivel fundamental, la vida marina afecta la naturaleza del planeta. Los organismos marinos, en su mayoría microorganismos, producen oxígeno y secuestran carbono. La vida marina, en parte, da forma y protege las costas, y algunos organismos marinos incluso ayudan a crear nuevas tierras (por ejemplo, los arrecifes de coral).
La mayoría de las formas de vida evolucionaron inicialmente en hábitats marinos. En volumen, los océanos proporcionan alrededor del 90% del espacio habitable del planeta. Los primeros vertebrados aparecieron en forma de peces, que vivían exclusivamente en el agua. Algunos de ellos evolucionaron hasta convertirse en anfibios, que pasan partes de sus vidas en el agua y otras en la tierra. Un grupo de anfibios evolucionó hasta convertirse en reptiles y mamíferos y algunos subconjuntos de cada uno regresaron al océano como serpientes marinas, tortugas marinas, focas, manatíes y ballenas. Formas vegetales como las algas marinas y otras algas crecen en el agua y son la base de algunos ecosistemas submarinos. El plancton constituye la base general de la cadena alimentaria oceánica, en particular el fitoplancton, que es un productor primario clave.
Los invertebrados marinos exhiben una amplia gama de modificaciones para sobrevivir en aguas poco oxigenadas, incluidos tubos de respiración como los sifones de los moluscos. Los peces tienen branquias en lugar de pulmones, aunque algunas especies de peces, como el pez pulmonado, tienen ambos. Los mamíferos marinos (por ejemplo, delfines, ballenas, nutrias y focas) necesitan salir a la superficie periódicamente para respirar aire.
A partir de 2023, se han documentado más de 242.000 especies marinas, y quizás aún quedan dos millones de especies marinas por documentar. Se describen una media de 2.332 nuevas especies al año.
Las especies marinas varían en tamaño desde microscópicas como el fitoplancton, que puede ser tan pequeño como 0,02 micrómetros, hasta enormes cetáceos como la ballena azul, el animal más grande conocido, que alcanza los 33 m (108 pies) de longitud. Se ha estimado que los microorganismos marinos, incluidos los protistas y las bacterias y sus virus asociados, constituyen alrededor del 70% o alrededor del 90% de la biomasa marina total. La vida marina se estudia científicamente tanto en biología marina como en oceanografía biológica. El término marino proviene del latín mare, que significa "mar" o "océano".
Agua

No hay vida sin agua. Ha sido descrito como el disolvente universal por su capacidad para disolver muchas sustancias, y como el disolvente de la vida. El agua es la única sustancia común que existe como sólido, líquido y gas en condiciones normales para la vida en la Tierra. El premio Nobel Albert Szent-Györgyi se refirió al agua como mater und Matrix: la madre y el útero de la vida.

La abundancia de agua superficial en la Tierra es una característica única en el Sistema Solar. La hidrosfera de la Tierra se compone principalmente de océanos, pero técnicamente incluye todas las superficies de agua del mundo, incluidos mares interiores, lagos, ríos y aguas subterráneas hasta una profundidad de 2.000 metros (6.600 pies). El lugar submarino más profundo es el abismo Challenger de la Fosa de las Marianas en el Océano Pacífico, con una profundidad de 10.900 metros (6,8 millas).
Convencionalmente, el planeta está dividido en cinco océanos separados, pero todos estos océanos se conectan en un solo océano mundial. La masa de este océano mundial es 1,35×10<span data-sort-value="7001180000000000000♠" 18 toneladas métricas o aproximadamente 1/4400 de la masa total de la Tierra. El océano mundial cubre un área de 3,618×108 km2 con una profundidad media de 3682 m, lo que da como resultado un volumen estimado de 1,332 ×109 km3. Si toda la superficie de la corteza terrestre estuviera a la misma altura que una esfera lisa, la profundidad del océano mundial resultante sería de unos 2,7 kilómetros (1,7 millas).

Aproximadamente el 97,5% del agua de la Tierra es salina; el 2,5% restante es agua dulce. La mayor parte del agua dulce (alrededor del 69%) está presente en forma de hielo en casquetes polares y glaciares. La salinidad promedio de los océanos de la Tierra es de aproximadamente 35 gramos (1,2 oz) de sal por kilogramo de agua de mar (3,5% de sal). La mayor parte de la sal del océano proviene de la erosión de las rocas terrestres. Algunas sales se liberan por la actividad volcánica o se extraen de rocas ígneas frías.
Los océanos también son un depósito de gases atmosféricos disueltos, que son esenciales para la supervivencia de muchas formas de vida acuática. El agua de mar tiene una influencia importante en el clima mundial, ya que los océanos actúan como una gran reserva de calor. Los cambios en la distribución de la temperatura oceánica pueden causar cambios climáticos significativos, como El Niño-Oscilación del Sur.

En total, el océano ocupa el 71 por ciento de la superficie mundial, con un promedio de casi 3,7 kilómetros (2,3 millas) de profundidad. En volumen, el océano proporciona alrededor del 90 por ciento del espacio habitable del planeta. El escritor de ciencia ficción Arthur C. Clarke ha señalado que sería más apropiado referirse al planeta Tierra como planeta Océano.
Sin embargo, el agua se encuentra en otras partes del Sistema Solar. Europa, una de las lunas que orbitan alrededor de Júpiter, es ligeramente más pequeña que la Luna de la Tierra. Existe una gran posibilidad de que exista un gran océano de agua salada debajo de su superficie de hielo. Se ha estimado que la corteza exterior de hielo sólido tiene entre 10 y 30 km (6 a 19 millas) de espesor y el océano líquido que se encuentra debajo tiene unos 100 km (60 millas) de profundidad. Esto haría que el océano de Europa duplicara el volumen del océano de la Tierra. Se ha especulado que el océano de Europa podría sustentar vida y podría ser capaz de sustentar microorganismos multicelulares si los respiraderos hidrotermales están activos en el fondo del océano. Encelado, una pequeña luna helada de Saturno, también tiene lo que parece ser un océano subterráneo que expulsa activamente agua caliente de la superficie de la luna.
Evolución
Duración de la vida | ||||||||||||||||||||||||||||||||||||||||||||||
−4500 — – — – 4.000 - 4000 – — – −3500 – — – −3000 – – — – −2500 — – — – −2000 - – — – −1500 — – — – 1000−1000 – — – .500 - 500 – — – 0 - | Agua Vida única Fotosíntesis Eukaryotes Vida multicelular Plantas Artropods Molluscs Flores Dinosaurios Mamíferos Aves Primados Hadean Archean Proterozoico Phanerozoic |
| ||||||||||||||||||||||||||||||||||||||||||||
(Hace millones de años) *Edades de hielo | ||||||||||||||||||||||||||||||||||||||||||||||
Desarrollo histórico
La Tierra tiene aproximadamente 4,54 mil millones de años. La evidencia indiscutible más antigua de vida en la Tierra data de hace al menos 3.500 millones de años, durante la era Eoarqueana, después de que una corteza geológica comenzara a solidificarse después del anterior Eón Hadeano fundido. Se han encontrado fósiles de estera microbiana en arenisca de 3.480 millones de años en Australia Occidental. Otra evidencia física temprana de una sustancia biogénica es el grafito en rocas metasedimentarias de 3.700 millones de años descubiertas en el oeste de Groenlandia, así como "restos de vida biótica" encontrado en rocas de 4.100 millones de años en Australia Occidental. Según uno de los investigadores, "si la vida surgió relativamente rápido en la Tierra... entonces podría ser común en el universo".
Todos los organismos de la Tierra descienden de un ancestro común o de un acervo genético ancestral. Se cree que la química altamente energética produjo una molécula autorreplicante hace unos 4 mil millones de años, y 500 millones de años después existió el último ancestro común de toda la vida. El consenso científico actual es que la compleja bioquímica que constituye la vida provino de reacciones químicas más simples. El comienzo de la vida puede haber incluido moléculas autorreplicantes como el ARN y el ensamblaje de células simples. En 2016, los científicos informaron sobre un conjunto de 355 genes del último ancestro común universal (LUCA) de toda la vida, incluidos los microorganismos, que viven en la Tierra.
Las especies actuales son una etapa en el proceso de evolución, siendo su diversidad el producto de una larga serie de eventos de especiación y extinción. La descendencia común de los organismos se dedujo primero de cuatro hechos simples sobre los organismos: primero, tienen distribuciones geográficas que no pueden explicarse mediante una adaptación local. En segundo lugar, la diversidad de la vida no es un conjunto de organismos únicos, sino organismos que comparten similitudes morfológicas. En tercer lugar, los rasgos vestigiales sin un propósito claro se parecen a los rasgos ancestrales funcionales y, finalmente, los organismos pueden clasificarse utilizando estas similitudes en una jerarquía de grupos anidados, similar a un árbol genealógico. Sin embargo, la investigación moderna ha sugerido que, debido a la transferencia horizontal de genes, este "árbol de la vida" Puede ser más complicado que un simple árbol ramificado, ya que algunos genes se han propagado de forma independiente entre especies lejanamente relacionadas.
Las especies del pasado también han dejado registros de su historia evolutiva. Los fósiles, junto con la anatomía comparada de los organismos actuales, constituyen el registro morfológico o anatómico. Al comparar las anatomías de especies modernas y extintas, los paleontólogos pueden inferir los linajes de esas especies. Sin embargo, este enfoque tiene más éxito en organismos que tenían partes duras del cuerpo, como caparazones, huesos o dientes. Además, como los procariotas como las bacterias y las arqueas comparten un conjunto limitado de morfologías comunes, sus fósiles no proporcionan información sobre su ascendencia.

Más recientemente, la evidencia de una ascendencia común proviene del estudio de las similitudes bioquímicas entre organismos. Por ejemplo, todas las células vivas utilizan el mismo conjunto básico de nucleótidos y aminoácidos. El desarrollo de la genética molecular ha revelado el registro de la evolución dejado en los organismos. Genomas: datación cuando las especies divergieron a través del reloj molecular producido por mutaciones. Por ejemplo, estas comparaciones de secuencias de ADN han revelado que los humanos y los chimpancés comparten el 98% de sus genomas y el análisis de las pocas áreas en las que difieren ayuda a arrojar luz sobre cuándo existió el ancestro común de estas especies.
Los procariotas habitaron la Tierra hace aproximadamente 3 a 4 mil millones de años. No se produjeron cambios obvios en la morfología o la organización celular en estos organismos durante los siguientes miles de millones de años. Las células eucariotas surgieron hace entre 1.600 y 2.700 millones de años. El siguiente cambio importante en la estructura celular se produjo cuando las bacterias fueron fagocitadas por células eucariotas, en una asociación cooperativa llamada endosimbiosis. Las bacterias engullidas y la célula huésped experimentaron luego una coevolución, y las bacterias evolucionaron hacia mitocondrias o hidrogenosomas. Otra absorción de organismos similares a las cianobacterias condujo a la formación de cloroplastos en algas y plantas.

La historia de la vida fue la de los eucariotas unicelulares, procariotas y arqueas hasta hace unos 610 millones de años, cuando comenzaron a aparecer organismos multicelulares en los océanos en el período Ediacárico. La evolución de la multicelularidad se produjo en múltiples eventos independientes, en organismos tan diversos como esponjas, algas pardas, cianobacterias, mohos mucilaginosos y mixobacterias. En 2016, los científicos informaron que, hace unos 800 millones de años, un cambio genético menor en una sola molécula llamada GK-PID pudo haber permitido que los organismos pasaran de ser un organismo unicelular a una de muchas células.
Poco después de la aparición de estos primeros organismos multicelulares, apareció una cantidad notable de diversidad biológica en un lapso de aproximadamente 10 millones de años, en un evento llamado explosión del Cámbrico. Aquí aparecieron en el registro fósil la mayoría de los tipos de animales modernos, así como linajes únicos que posteriormente se extinguieron. Se han propuesto varios desencadenantes de la explosión del Cámbrico, incluida la acumulación de oxígeno en la atmósfera a partir de la fotosíntesis.
Hace unos 500 millones de años, las plantas y los hongos comenzaron a colonizar la tierra. La evidencia de la aparición de las primeras plantas terrestres se produce en el Ordovícico, alrededor de hace 450 millones de años, en forma de esporas fósiles. Las plantas terrestres comenzaron a diversificarse a finales del Silúrico, hace alrededor de hace 430 millones de años. A la colonización de la tierra por plantas pronto le siguieron artrópodos y otros animales. Los insectos tuvieron especial éxito y aún hoy constituyen la mayoría de las especies animales. Los anfibios aparecieron por primera vez hace unos 364 millones de años, seguidos por los primeros amniotas y las aves hace unos 155 millones de años (ambos de linajes similares a los de los "reptiles"), los mamíferos hace unos 129 millones de años, los homínidos hace unos 10 millones de años y Los humanos modernos hace unos 250.000 años. Sin embargo, a pesar de la evolución de estos animales grandes, organismos más pequeños similares a los tipos que evolucionaron al principio de este proceso continúan teniendo mucho éxito y dominando la Tierra, siendo la mayoría de la biomasa y las especies procariotas.
Las estimaciones sobre el número de especies actuales en la Tierra oscilan entre 10 y 14 millones, de las cuales alrededor de 1,2 millones han sido documentadas y más del 86 por ciento aún no han sido descritas.
Microorganismos


Los microorganismos constituyen aproximadamente el 70% de la biomasa marina. Un microorganismo, o microbio, es un organismo microscópico demasiado pequeño para ser reconocido a simple vista. Puede ser unicelular o multicelular. Los microorganismos son diversos e incluyen todas las bacterias y arqueas, la mayoría de los protozoos, como las algas, los hongos y ciertos animales microscópicos, como los rotíferos.
Muchos animales y plantas macroscópicos tienen etapas juveniles microscópicas. Algunos microbiólogos también clasifican los virus (y viroides) como microorganismos, pero otros los consideran no vivos.
Los microorganismos son cruciales para el reciclaje de nutrientes en los ecosistemas, ya que actúan como descomponedores. Algunos microorganismos son patógenos y causan enfermedades e incluso la muerte en plantas y animales. Como habitantes del entorno más grande de la Tierra, los sistemas marinos microbianos impulsan cambios en todos los sistemas globales. Los microbios son responsables de prácticamente toda la fotosíntesis que ocurre en el océano, así como del ciclo del carbono, nitrógeno, fósforo, otros nutrientes y oligoelementos.

| Microorganismos marinos |
| |||||||||||||||||||||||||||
_-_Stephanopyxis_sp._-_630x_(14117161428).jpg)

La vida microscópica submarina es diversa y aún no se comprende bien, como por ejemplo el papel de los virus en los ecosistemas marinos. La mayoría de los virus marinos son bacteriófagos, que son inofensivos para las plantas y los animales, pero son esenciales para la regulación de los ecosistemas de agua dulce y salada. Infectan y destruyen bacterias en comunidades microbianas acuáticas y son el mecanismo más importante de reciclaje de carbono en el medio marino. Las moléculas orgánicas liberadas por las células bacterianas muertas estimulan el crecimiento de bacterias y algas frescas. La actividad viral también puede contribuir a la bomba biológica, el proceso mediante el cual el carbono se secuestra en las profundidades del océano.


Una corriente de microorganismos transportados por el aire circula alrededor del planeta por encima de los sistemas climáticos pero por debajo de las rutas aéreas comerciales. Algunos microorganismos peripatéticos son arrastrados por las tormentas de polvo terrestres, pero la mayoría se origina a partir de microorganismos marinos en la espuma del mar. En 2018, los científicos informaron que diariamente se depositan cientos de millones de virus y decenas de millones de bacterias en cada metro cuadrado del planeta.
Los organismos microscópicos viven en toda la biosfera. La masa de microorganismos procariotas (que incluye bacterias y arqueas, pero no los microorganismos eucariotas nucleados) puede alcanzar los 0,8 billones de toneladas de carbono (de la masa total de la biosfera, estimada entre 1 y 4 billones de toneladas). Se han encontrado microbios marinos barófilos unicelulares a una profundidad de 10.900 m (35.800 pies) en la Fosa de las Marianas, el lugar más profundo de los océanos de la Tierra. Los microorganismos viven dentro de rocas a 580 m (1900 pies) debajo del fondo del mar, a 2590 m (8500 pies) de océano frente a la costa del noroeste de Estados Unidos, así como a 2400 m (7900 pies; 1,5 millas) debajo del lecho marino frente a Japón. La temperatura más alta conocida a la que puede existir vida microbiana es 122 °C (252 °F) (Methanopyrus kandleri). En 2014, los científicos confirmaron la existencia de microorganismos que viven a 800 m (2600 pies) debajo del hielo de la Antártida. Según un investigador, "se pueden encontrar microbios en todas partes: son extremadamente adaptables a las condiciones y sobreviven dondequiera que estén".
Virus marinos
Los virus son pequeños agentes infecciosos que no tienen metabolismo propio y sólo pueden replicarse dentro de las células vivas de otros organismos. Los virus pueden infectar todo tipo de formas de vida, desde animales y plantas hasta microorganismos, incluidas bacterias y arqueas. El tamaño lineal de un virus promedio es aproximadamente una centésima parte del de una bacteria promedio. La mayoría de los virus no pueden verse con un microscopio óptico, por lo que en su lugar se utilizan microscopios electrónicos.
Los virus se encuentran donde hay vida y probablemente han existido desde que las células vivas evolucionaron primero. El origen de los virus no está claro porque no forman fósiles, por lo que se han utilizado técnicas moleculares para comparar el ADN o ARN de virus y son un medio útil de investigar cómo se presentan.
Ahora se reconoce que los virus son antiguos y tienen orígenes anteriores a la divergencia de la vida en los tres dominios. Pero los orígenes de los virus en la historia evolutiva de la vida no están claros: algunos pueden haber evolucionado a partir de plásmidos (fragmentos de ADN que pueden moverse entre células), mientras que otros pueden haber evolucionado a partir de bacterias. En la evolución, los virus son un medio importante de transferencia horizontal de genes, lo que aumenta la diversidad genética.



Las opiniones difieren sobre si los virus son una forma de vida o estructuras orgánicas que interactúan con organismos vivos. Algunos los consideran una forma de vida porque portan material genético, se reproducen creando múltiples copias de sí mismos mediante autoensamblaje y evolucionan mediante selección natural. Sin embargo, carecen de características clave, como una estructura celular que generalmente se considera necesaria para contar como vida. Debido a que poseen algunas de esas cualidades, pero no todas, los virus han sido descritos como replicadores y "organismos al borde de la vida".

Los bacteriófagos, a menudo llamados simplemente fagos, son virus que parasitan bacterias y arqueas. Los fagos marinos parasitan bacterias y arqueas marinas, como las cianobacterias. Son un grupo común y diverso de virus y son la entidad biológica más abundante en ambientes marinos, porque sus huéspedes, las bacterias, suelen ser la vida celular numéricamente dominante en el mar. En general, hay entre 1 y 10 millones de virus en cada ml de agua de mar, o aproximadamente diez veces más virus de ADN de doble hebra que organismos celulares, aunque las estimaciones de la abundancia viral en el agua de mar pueden variar en un amplio rango. Los bacteriófagos con cola parecen dominar los ecosistemas marinos en número y diversidad de organismos. También se sabe que los bacteriófagos pertenecientes a las familias Corticoviridae, Inoviridae y Microviridae infectan diversas bacterias marinas.
Los microorganismos representan alrededor del 70% de la biomasa marina. Se estima que los virus matan el 20% de esta biomasa cada día y que hay 15 veces más virus en los océanos ya que hay bacterias y arqueas. Los virus son los principales agentes responsables de la rápida destrucción de las floraciones de algas dañinas, que a menudo matan otra vida marina. El número de virus en los océanos disminuye aún más en el mar y más en el agua, donde hay menos organismos anfitriones.
También hay virus de arqueas que se replican dentro de arqueas: se trata de virus de ADN de doble cadena con formas inusuales y, a veces, únicas. Estos virus se han estudiado con mayor detalle en las arqueas termófilas, particularmente en los órdenes Sulfolobales y Thermoproteales.
Los virus son un medio natural importante para transferir genes entre diferentes especies, lo que aumenta la diversidad genética e impulsa la evolución. Se cree que los virus desempeñaron un papel central en la evolución temprana, antes de la diversificación de bacterias, arqueas y eucariotas, en la época del último ancestro común universal de la vida en la Tierra. Los virus siguen siendo uno de los mayores reservorios de diversidad genética inexplorada en la Tierra.
Bacterias marinas


Las bacterias constituyen un gran dominio de microorganismos procarióticos. Las bacterias, que suelen tener una longitud de unos pocos micrómetros, tienen diversas formas, que van desde esferas hasta varillas y espirales. Las bacterias estuvieron entre las primeras formas de vida que aparecieron en la Tierra y están presentes en la mayoría de sus hábitats. Las bacterias habitan en el suelo, el agua, las aguas termales ácidas, los desechos radiactivos y las porciones profundas de la corteza terrestre. Las bacterias también viven en relaciones simbióticas y parasitarias con plantas y animales.
Las bacterias, antes consideradas plantas que constituían la clase esquizomicetos, ahora se clasifican como procariotas. A diferencia de las células de animales y otros eucariotas, las células bacterianas no contienen núcleo y rara vez albergan orgánulos unidos a membranas. Aunque el término bacteria tradicionalmente incluía a todos los procariotas, la clasificación científica cambió después del descubrimiento en la década de 1990 de que los procariotas consisten en dos grupos muy diferentes de organismos que evolucionaron a partir de un antiguo ancestro común. Estos dominios evolutivos se denominan Bacteria y Archaea.
Los antepasados de las bacterias modernas fueron microorganismos unicelulares que fueron las primeras formas de vida que aparecieron en la Tierra, hace unos 4 mil millones de años. Durante unos 3 mil millones de años, la mayoría de los organismos eran microscópicos y las bacterias y arqueas eran las formas de vida dominantes. Aunque existen fósiles de bacterias, como los estromatolitos, su falta de morfología distintiva impide que se utilicen para examinar la historia de la evolución bacteriana o para fechar el momento de origen de una especie bacteriana en particular. Sin embargo, se pueden utilizar secuencias genéticas para reconstruir la filogenia bacteriana, y estos estudios indican que las bacterias divergieron primero del linaje arqueal/eucariota. Las bacterias también estuvieron implicadas en la segunda gran divergencia evolutiva, la de las arqueas y los eucariotas. Aquí, los eucariotas resultaron de la entrada de bacterias antiguas en asociaciones endosimbióticas con los antepasados de las células eucariotas, que posiblemente estaban relacionadas con las Archaea. Esto implicó la absorción por células protoeucariotas de simbiontes alfaproteobacterianos para formar mitocondrias o hidrogenosomas, que todavía se encuentran en todos los Eukarya conocidos. Más tarde, algunos eucariotas que ya contenían mitocondrias también engulleron organismos similares a las cianobacterias. Esto llevó a la formación de cloroplastos en algas y plantas. También hay algunas algas que se originaron a partir de eventos endosimbióticos incluso posteriores. Aquí, los eucariotas engulleron un alga eucariota que se convirtió en un alga de "segunda generación" plástido. Esto se conoce como endosimbiosis secundaria.
 El marino Thiomargarita namibiensis, la bacteria más grande conocida
El marino Thiomargarita namibiensis, la bacteria más grande conocida Las floraciones de Cyanobacteria pueden contener cyanotoxinas letales.
Las floraciones de Cyanobacteria pueden contener cyanotoxinas letales.![The chloroplasts of glaucophytes have a peptidoglycan layer, evidence suggesting their endosymbiotic origin from cyanobacteria.[132]](https://upload.wikimedia.org/wikipedia/commons/thumb/c/c9/Glaucocystis_sp.jpg/280px-Glaucocystis_sp.jpg) Los cloroplastos de glaucofitos tienen una capa peptidoglycan, evidencia que sugiere su origen endosymbiótico de cianobacteria.
Los cloroplastos de glaucofitos tienen una capa peptidoglycan, evidencia que sugiere su origen endosymbiótico de cianobacteria. Las bacterias pueden ser beneficiosas. Este gusano Pompeya, un extremofilo encontrado sólo en los respiraderos hidrotermales, tiene una cubierta protectora de bacterias.
Las bacterias pueden ser beneficiosas. Este gusano Pompeya, un extremofilo encontrado sólo en los respiraderos hidrotermales, tiene una cubierta protectora de bacterias.


![The chloroplasts of glaucophytes have a peptidoglycan layer, evidence suggesting their endosymbiotic origin from cyanobacteria.[132]](https://en.wikipedia.org/wiki/File:Glaucocystis_sp.jpg)

La bacteria más grande conocida, la Thiomargarita namibiensis marina, puede ser visible a simple vista y en ocasiones alcanza 0,75 mm (750 μm).
Arqueas marinas

Las arqueas (en griego, antiguas) constituyen un dominio y reino de microorganismos unicelulares. Estos microbios son procariotas, lo que significa que no tienen núcleo celular ni ningún otro orgánulo unido a una membrana en sus células.
Las arqueas fueron inicialmente clasificadas como bacterias, pero esta clasificación está desactualizada. Las células de arqueas tienen propiedades únicas que las separan de los otros dos dominios de la vida, bacterias y eucariotas. Las Archaea se dividen a su vez en múltiples filos reconocidos. La clasificación es difícil porque la mayoría no han sido aislados en laboratorio y sólo han sido detectados mediante análisis de sus ácidos nucleicos en muestras de su entorno.
Las arqueas y las bacterias son generalmente similares en tamaño y forma, aunque algunas arqueas tienen formas muy extrañas, como las células planas y cuadradas de Haloquadratum walsbyi. A pesar de esta similitud morfológica con las bacterias, las arqueas poseen genes y varias vías metabólicas que están más estrechamente relacionadas con las de los eucariotas, en particular las enzimas implicadas en la transcripción y la traducción. Otros aspectos de la bioquímica de las arqueas son únicos, como su dependencia de los éter-lípidos en sus membranas celulares, como los arqueoles. Las arqueas utilizan más fuentes de energía que los eucariotas: desde compuestos orgánicos, como azúcares, hasta amoníaco, iones metálicos o incluso gas hidrógeno. Las arqueas tolerantes a la sal (las Haloarchaea) utilizan la luz solar como fuente de energía, y otras especies de arqueas fijan carbono; sin embargo, a diferencia de las plantas y las cianobacterias, ninguna especie conocida de arquea hace ambas cosas. Archaea se reproduce asexualmente por fisión binaria, fragmentación o gemación; a diferencia de las bacterias y los eucariotas, ninguna especie conocida forma esporas.
Las arqueas son particularmente numerosas en los océanos, y las arqueas del plancton pueden ser uno de los grupos de organismos más abundantes del planeta. Las arqueas son una parte importante de la vida en la Tierra y pueden desempeñar funciones tanto en el ciclo del carbono como en el ciclo del nitrógeno.
 La halobacteria, que se encuentra en el agua cerca de saturada de sal, ahora son reconocidas como arqueas.
La halobacteria, que se encuentra en el agua cerca de saturada de sal, ahora son reconocidas como arqueas. Células planas en forma cuadrada de la arquea Haloquadratum walsbyi
Células planas en forma cuadrada de la arquea Haloquadratum walsbyi Methanosarcina barkeri, una arquea marina que produce metano
Methanosarcina barkeri, una arquea marina que produce metano Termófilos, como Pyrolobus fumariiSobrevive mucho más de 100 °C.
Termófilos, como Pyrolobus fumariiSobrevive mucho más de 100 °C. Dibujo de otro termófilo marino, Pyrococcus furiosus
Dibujo de otro termófilo marino, Pyrococcus furiosus





Protistas marinos
Los protistas son eucariotas que no pueden clasificarse como plantas, hongos o animales. Suelen ser unicelulares y microscópicos. La vida se originó como procariotas unicelulares (bacterias y arqueas) y luego evolucionó hacia eucariotas más complejos. Los eucariotas son las formas de vida más desarrolladas conocidas como plantas, animales, hongos y protistas. El término protista se empezó a utilizar históricamente como un término de conveniencia para eucariotas que no pueden clasificarse estrictamente como plantas, animales u hongos. No forman parte de la cladística moderna porque son parafiléticos (carecen de un ancestro común). Los protistas se pueden dividir en cuatro grupos dependiendo de si su nutrición es similar a una planta, a un animal, a un hongo o una mezcla de estos.
Protistas según cómo consiguen comida | |||||||
|---|---|---|---|---|---|---|---|
| Tipo de protista | Descripción | Ejemplo | Otros ejemplos | ||||
| Como las plantas | Algae (véase infra) | Protistas autotróficos que hacen su propio alimento sin necesidad de consumir otros organismos, generalmente usando fotosíntesis |  | Alga roja, Cyanidium sp. | Algas verdes, algas marrones, diatomeas y algunos dinoflagelados. Los protistas similares a las plantas son componentes importantes del fitoplancton discutido a continuación. | ||
| Como animales | Protozoans | Protistas heterotróficos que consumen sus alimentos otros organismos |  | Protista radilar dibujado por Haeckel | Foraminiferans, and some marine amoebae, ciliates and flagellates. | ||
| Fungus-como | Moldes de deslizamiento y redes slime | Protistas saprotróficos que obtienen su alimento de los restos de organismos que han descompuesto y descompuesto |  | Redes slime marinas forman redes laberínticas de tubos en los que las ameba sin pseudopodos pueden viajar | Lichen marino | ||
| Mixotropos | Varios | Protistas mixotróficos y osmotróficos que obtienen su comida de una combinación de lo anterior | _(cropped).jpg) | Euglena mutabilis, un bandellate fotosintético | Muchos mixotropos marinos se encuentran entre los protistas, incluyendo entre los ciliates, Rhizaria y los dinoflagelados | ||


Los protistas son organismos muy diversos actualmente organizados en 18 filos, pero no son fáciles de clasificar. Los estudios han demostrado que existe una gran diversidad de protistas en los océanos, los respiraderos marinos profundos y los sedimentos de los ríos, lo que sugiere que aún no se ha descubierto una gran cantidad de comunidades microbianas eucariotas. Ha habido poca investigación sobre los protistas mixotróficos, pero estudios recientes en ambientes marinos encontraron que las protestas mixotróficas contribuyen con una parte significativa de la biomasa de protistas.
- Protistas monocelulares y microscópicos
![Diatoms are a major algae group generating about 20% of world oxygen production.[147]](https://upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg) Los átomos son un grupo de algas importante que genera alrededor del 20% de la producción mundial de oxígeno.
Los átomos son un grupo de algas importante que genera alrededor del 20% de la producción mundial de oxígeno.![Diatoms have glass like cell walls made of silica and called frustules.[148]](https://upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg) Los diatomeas tienen vidrio como paredes celulares hechas de sílice y llamadas frustules.
Los diatomeas tienen vidrio como paredes celulares hechas de sílice y llamadas frustules. Fossil diatom frustule de 32 a 40 mia
Fossil diatom frustule de 32 a 40 mia Radiolarian
Radiolarian Alga única, Gephyrocapsa oceanica
Alga única, Gephyrocapsa oceanica Dos dinoflagelados
Dos dinoflagelados Zooxanthellae es una alga fotosintética que vive dentro de hosts como coral.
Zooxanthellae es una alga fotosintética que vive dentro de hosts como coral. Un ciliato monocelular con zooclorella verde que vive dentro endosimbióticamente.
Un ciliato monocelular con zooclorella verde que vive dentro endosimbióticamente. Euglenoid
Euglenoid Este ciliato está digeriendo cianobacteria. El citotomoma o la boca está en la parte inferior derecha.
Este ciliato está digeriendo cianobacteria. El citotomoma o la boca está en la parte inferior derecha.
![Diatoms are a major algae group generating about 20% of world oxygen production.[147]](https://en.wikipedia.org/wiki/File:Diatoms_through_the_microscope.jpg)
![Diatoms have glass like cell walls made of silica and called frustules.[148]](https://en.wikipedia.org/wiki/File:Diatom_algae_Amphora_sp.jpg)

.jpg)
.jpg)





A diferencia de las células procariotas, las células eucariotas están muy organizadas. Las plantas, los animales y los hongos suelen ser multicelulares y típicamente macroscópicos. La mayoría de los protistas son unicelulares y microscópicos. Pero hay excepciones. Algunos protistas marinos unicelulares son macroscópicos. Algunos mohos del limo marino tienen ciclos de vida únicos que implican el cambio entre formas unicelulares, coloniales y multicelulares. Otros protistas marinos no son unicelulares ni microscópicos, como las algas.
- Protistas macroscópicos (ver también macroalgas unicelares →)
 La ameba gigante monocelada tiene hasta 1000 núcleos y alcanza longitudes de 5 mm (0.20 en).
La ameba gigante monocelada tiene hasta 1000 núcleos y alcanza longitudes de 5 mm (0.20 en).![Gromia sphaerica is a large spherical testate amoeba which makes mud trails. Its diameter is up to 3.8 cm (1.5 in).[150]](https://upload.wikimedia.org/wikipedia/commons/thumb/9/99/Gromia_in_situ_closeup.png/266px-Gromia_in_situ_closeup.png) Gromia sphaerica es una gran testate ameba esférica que hace senderos de barro. Su diámetro es de hasta 3.8 cm (1.5 pulgadas).
Gromia sphaerica es una gran testate ameba esférica que hace senderos de barro. Su diámetro es de hasta 3.8 cm (1.5 pulgadas). Spiculosiphon oceana, un foraminiferan unicellular con una apariencia y estilo de vida que imita una esponja, crece a 5 cm de largo.
Spiculosiphon oceana, un foraminiferan unicellular con una apariencia y estilo de vida que imita una esponja, crece a 5 cm de largo.![The xenophyophore, another single-celled foraminiferan, lives in abyssal zones. It has a giant shell up to 20 cm (7.9 in) across.[151]](https://upload.wikimedia.org/wikipedia/commons/thumb/5/5d/Xenophyophore.jpg/292px-Xenophyophore.jpg) El xenophyophore, otro foraminiferano monocelular, vive en zonas abisales. Tiene una cáscara gigante de hasta 20 cm (7.9 pulgadas) de ancho.
El xenophyophore, otro foraminiferano monocelular, vive en zonas abisales. Tiene una cáscara gigante de hasta 20 cm (7.9 pulgadas) de ancho. El kelp gigante, una alga marrón, no es una verdadera planta, sin embargo es multicelular y puede crecer a 50m.
El kelp gigante, una alga marrón, no es una verdadera planta, sin embargo es multicelular y puede crecer a 50m.

![Gromia sphaerica is a large spherical testate amoeba which makes mud trails. Its diameter is up to 3.8 cm (1.5 in).[150]](https://en.wikipedia.org/wiki/File:Gromia_in_situ_closeup.png)

![The xenophyophore, another single-celled foraminiferan, lives in abyssal zones. It has a giant shell up to 20 cm (7.9 in) across.[151]](https://en.wikipedia.org/wiki/File:Xenophyophore.jpg)

Los protistas han sido descritos como una bolsa de sorpresas taxonómica donde se puede colocar cualquier cosa que no encaje en uno de los principales reinos biológicos. Algunos autores modernos prefieren excluir los organismos multicelulares de la definición tradicional de protista, restringiendo los protistas a organismos unicelulares. Esta definición más restringida excluye las algas y los mohos mucilaginosos.
Microanimales marinos
Como juveniles, los animales se desarrollan a partir de etapas microscópicas, que pueden incluir esporas, huevos y larvas. Al menos un grupo de animales microscópicos, el cnidario parásito Myxozoa, es unicelular en su forma adulta e incluye especies marinas. Otros microanimales marinos adultos son multicelulares. Los artrópodos adultos microscópicos se encuentran más comúnmente en el interior, en agua dulce, pero también hay especies marinas. Los crustáceos marinos adultos microscópicos incluyen algunos copépodos, cladóceros y tardígrados (osos de agua). Algunos nematodos y rotíferos marinos también son demasiado pequeños para ser reconocidos a simple vista, al igual que muchos loricíferos, incluidas las especies anaeróbicas recientemente descubiertas que pasan su vida en un ambiente anóxico. Los copépodos contribuyen más a la productividad secundaria y al sumidero de carbono de los océanos del mundo que cualquier otro grupo de organismos. Si bien normalmente no se considera que los ácaros sean organismos marinos, la mayoría de las especies de la familia Halacaridae viven en el mar.
- Microanimales marinos
 Más de 10.000 especies marinas son coppodos, pequeños, a menudo microscópicos crustáceos
Más de 10.000 especies marinas son coppodos, pequeños, a menudo microscópicos crustáceos Foto de campo oscuro de un gastrotrico, un animal parecido a gusano que vive entre partículas de sedimento
Foto de campo oscuro de un gastrotrico, un animal parecido a gusano que vive entre partículas de sedimento Armoured Pliciloricus enigmaticus, aproximadamente 0,2 mm de largo, vivir en espacios entre la grava marina.
Armoured Pliciloricus enigmaticus, aproximadamente 0,2 mm de largo, vivir en espacios entre la grava marina. Dibujo de un tardigrado (oso de agua) sobre un grano de arena
Dibujo de un tardigrado (oso de agua) sobre un grano de arena Los rotifers, generalmente 0.1–0.5 mm de largo, pueden parecer protistas pero tienen muchas células y pertenece a la Animalia.
Los rotifers, generalmente 0.1–0.5 mm de largo, pueden parecer protistas pero tienen muchas células y pertenece a la Animalia.




_-_160x_(13402418244).jpg)
Hongos


Se conocen más de 1500 especies de hongos en ambientes marinos. Estos son parásitos de algas o animales marinos, o son saprobios que se alimentan de materia orgánica muerta de algas, corales, quistes de protozoos, pastos marinos, madera y otros sustratos. Las esporas de muchas especies tienen apéndices especiales que facilitan la unión al sustrato. Los hongos marinos también se pueden encontrar en la espuma del mar y alrededor de las zonas hidrotermales del océano. Los hongos marinos producen una amplia gama de metabolitos secundarios inusuales.
El micoplancton es miembro saprotrópico de las comunidades de plancton de los ecosistemas marinos y de agua dulce. Están compuestos por hongos filamentosos de vida libre y levaduras asociadas a partículas planctónicas o fitoplancton. Al igual que el bacterioplancton, estos hongos acuáticos desempeñan un papel importante en la mineralización heterótrofa y el ciclo de nutrientes. El micoplancton puede tener hasta 20 mm de diámetro y más de 50 mm de longitud.
Un mililitro típico de agua de mar contiene alrededor de 103 a 104 células fúngicas. Este número es mayor en los ecosistemas costeros y estuarios debido a la escorrentía nutricional de las comunidades terrestres. Una mayor diversidad de micoplancton se encuentra alrededor de las costas y en aguas superficiales hasta los 1.000 metros, con un perfil vertical que depende de la abundancia de fitoplancton. Este perfil cambia entre estaciones debido a cambios en la disponibilidad de nutrientes. Los hongos marinos sobreviven en un ambiente constantemente deficiente en oxígeno y, por lo tanto, dependen de la difusión de oxígeno mediante turbulencias y del oxígeno generado por organismos fotosintéticos.
Los hongos marinos se pueden clasificar en:
- hongos inferiores - adaptados a hábitats marinos ( hongos zoosporicos, incluyendo mastigomycetes: oomycetes y chytridiomycetes)
- hongos superiores - filamentosos, modificados al estilo de vida planctónico (hifomáceos, ascomicetos, basidiomycetes). La mayoría de las especies de micoplancton son hongos más altos.
Los líquenes son asociaciones mutualistas entre un hongo, generalmente un ascomiceto, y un alga o una cianobacteria. Varios líquenes se encuentran en ambientes marinos. Muchos más ocurren en la zona de salpicadura, donde ocupan diferentes zonas verticales dependiendo de su tolerancia a la inmersión. Algunos líquenes viven mucho tiempo; una especie ha sido datada en 8.600 años. Sin embargo, su esperanza de vida es difícil de medir porque lo que define al mismo liquen no es preciso. Los líquenes crecen rompiendo vegetativamente un trozo, que puede definirse o no como el mismo liquen, y dos líquenes de diferentes edades pueden fusionarse, planteando la cuestión de si se trata del mismo liquen. El caracol marino Littoraria irrorata daña las plantas de Spartina en las marismas donde vive, lo que permite que las esporas de hongos ascomicetos intermareales colonicen la planta. Luego, el caracol se come el crecimiento del hongo con preferencia a la hierba misma.
Según los registros fósiles, los hongos se remontan a finales de la era Proterozoica, hace entre 900 y 570 millones de años. En China se han descubierto líquenes marinos fósiles de 600 millones de años de antigüedad. Se ha planteado la hipótesis de que el micoplancton evolucionó a partir de hongos terrestres, probablemente en la era Paleozoica (hace 390 millones de años).
Origen de los animales

Los primeros animales fueron los invertebrados marinos, es decir, los vertebrados llegaron después. Los animales son eucariotas multicelulares y se distinguen de las plantas, algas y hongos por carecer de paredes celulares. Los invertebrados marinos son animales que habitan en un ambiente marino aparte de los miembros vertebrados del filo cordados; Los invertebrados carecen de columna vertebral. Algunos han desarrollado un caparazón o un exoesqueleto duro.
Los primeros fósiles de animales pueden pertenecer al género Dickinsonia, hace entre 571 y 541 millones de años. Los Dickinsonia individuales suelen parecerse a un óvalo acanalado con simetría bilateral. Siguieron creciendo hasta que quedaron cubiertos de sedimento o murieron de alguna otra manera, y pasaron la mayor parte de sus vidas con sus cuerpos firmemente anclados al sedimento. Actualmente se desconocen sus afinidades taxonómicas, pero su modo de crecimiento es consistente con una afinidad bilateral.
Aparte de Dickinsonia, los primeros fósiles de animales ampliamente aceptados son los cnidarios de aspecto bastante moderno (el grupo que incluye corales, medusas, anémonas de mar e hidras), posiblemente de alrededor de 580 Ma La biota de Ediacara, que floreció durante los últimos 40 millones de años antes del inicio del Cámbrico, fueron los primeros animales. más de unos pocos centímetros de largo. Como Dickinsonia, muchos eran planos con una manta "acolchada" apariencia, y parecía tan extraño que hubo una propuesta para clasificarlos como un reino separado, Vendozoa. Otros, sin embargo, han sido interpretados como moluscos primitivos (Kimberella), equinodermos (Arkarua) y artrópodos (Spriggina, Parvancorina). Todavía hay debate sobre la clasificación de estos especímenes, principalmente porque las características de diagnóstico que permiten a los taxónomos clasificar organismos más recientes, como las similitudes con organismos vivos, generalmente están ausentes en los ediacaranos. Sin embargo, parece haber pocas dudas de que Kimberella era al menos un animal bilateral triploblástico, es decir, un animal significativamente más complejo que los cnidarios.
La fauna de conchas pequeñas es una colección muy variada de fósiles encontrada entre los períodos Ediacárico Tardío y Cámbrico Medio. El más antiguo, Cloudina, muestra signos de defensa exitosa contra la depredación y puede indicar el inicio de una carrera armamentista evolutiva. Es casi seguro que algunas pequeñas conchas del Cámbrico temprano pertenecían a moluscos, mientras que los propietarios de algunas "placas de armadura" Halkieria y Microdictyon fueron finalmente identificados cuando se encontraron especímenes más completos en lagerstätten del Cámbrico que conservaban animales de cuerpo blando.
Planes corporales y filos

Los invertebrados se agrupan en diferentes filos. De manera informal, se puede pensar en los filos como una forma de agrupar organismos de acuerdo con su plan corporal. Un plan corporal se refiere a un plano que describe la forma o morfología de un organismo, como su simetría, segmentación y la disposición de sus apéndices. La idea de los planes corporales se originó en los vertebrados, que se agrupaban en un solo filo. Pero el plan corporal de los vertebrados es sólo uno de muchos, y los invertebrados constan de muchos filos o planes corporales. La historia del descubrimiento de los planes corporales puede verse como un movimiento desde una visión del mundo centrada en los vertebrados a ver a los vertebrados como un plan corporal entre muchos. Entre los zoólogos pioneros, Linneo identificó dos planos corporales fuera de los vertebrados; Cuvier identificó tres; y Haeckel tenía cuatro, así como el Protista con ocho más, para un total de doce. En comparación, el número de filos reconocidos por los zoólogos modernos ha aumentado a 35.


Históricamente se pensaba que los planes corporales habían evolucionado rápidamente durante la explosión del Cámbrico, pero una comprensión más matizada de la evolución animal sugiere un desarrollo gradual de los planes corporales a lo largo del Paleozoico temprano y más allá. De manera más general, un filo se puede definir de dos maneras: como se describió anteriormente, como un grupo de organismos con un cierto grado de similitud morfológica o de desarrollo (la definición fenética), o un grupo de organismos con un cierto grado de relación evolutiva (la definición filogenética). definición).
En la década de 1970 ya hubo un debate sobre si la aparición de los filos modernos fue “explosiva” o no. o gradual pero oculta por la escasez de fósiles de animales precámbricos. Un nuevo análisis de los fósiles de Burgess Shale lagerstätte aumentó el interés en el tema cuando reveló animales, como Opabinia, que no encajaban en ningún filo conocido. En ese momento, esto se interpretó como evidencia de que los filos modernos habían evolucionado muy rápidamente en la explosión del Cámbrico y que las "extrañas maravillas" de Burgess Shale no existían. demostró que el Cámbrico Inferior fue un período excepcionalmente experimental de evolución animal. Los descubrimientos posteriores de animales similares y el desarrollo de nuevos enfoques teóricos llevaron a la conclusión de que muchas de las "extrañas maravillas" eran "tías" o "primos" de grupos modernos: por ejemplo, que Opabinia era miembro de los lobopodos, un grupo que incluye a los antepasados de los artrópodos, y que puede haber estado estrechamente relacionado con los tardígrados modernos. Sin embargo, todavía hay mucho debate sobre si la explosión del Cámbrico fue realmente explosiva y, de ser así, cómo y por qué ocurrió y por qué parece única en la historia de los animales.
Los primeros animales
Los animales con ramificaciones más profundas (los primeros animales que aparecieron durante la evolución) son organismos marinos no vertebrados. Los primeros filos animales son Porifera, Ctenophora, Placozoa y Cnidaria. Ningún miembro de estos clados exhibe planes corporales con simetría bilateral.
| ||||||||||||||||||||||||||||||||||
.jpg)

Esponjas marinas
.jpg)
Las esponjas son animales del filo Porifera (del latín moderno para portar poros). Son organismos multicelulares que tienen cuerpos llenos de poros y canales que permiten que el agua circule a través de ellos, y consisten en mesohilo gelatinoso intercalado entre dos finas capas de células. Tienen células no especializadas que pueden transformarse en otros tipos y que a menudo migran entre las capas celulares principales y el mesohilo en el proceso. Las esponjas no tienen sistema nervioso, digestivo ni circulatorio. En cambio, la mayoría depende de mantener un flujo constante de agua a través de sus cuerpos para obtener alimentos y oxígeno y eliminar los desechos.
Las esponjas son similares a otros animales en que son multicelulares, heterótrofas, carecen de paredes celulares y producen espermatozoides. A diferencia de otros animales, carecen de verdaderos tejidos y órganos y no tienen simetría corporal. Las formas de sus cuerpos están adaptadas para lograr la máxima eficiencia del flujo de agua a través de la cavidad central, donde deposita los nutrientes, y sale por un orificio llamado ósculo. Muchas esponjas tienen esqueletos internos de espongina y/o espículas de carbonato cálcico o dióxido de silicio. Todas las esponjas son animales acuáticos sésiles. Aunque hay especies de agua dulce, la gran mayoría son especies marinas (de agua salada), que van desde zonas de marea hasta profundidades que superan los 8.800 m (5,5 millas). Algunas esponjas viven hasta edades avanzadas; Hay pruebas de que la esponja de cristal de aguas profundas Monorhaphis chuni vivió unos 11.000 años.
Si bien la mayoría de las aproximadamente 5.000 a 10.000 especies conocidas se alimentan de bacterias y otras partículas de alimentos en el agua, algunas albergan microorganismos fotosintetizadores como endosimbiontes y estas alianzas a menudo producen más alimentos y oxígeno de los que consumen. Unas pocas especies de esponjas que viven en ambientes pobres en alimentos se han convertido en carnívoros que se alimentan principalmente de pequeños crustáceos.
 Biodiversidad esponja. Hay cuatro especies de esponjas en esta foto.
Biodiversidad esponja. Hay cuatro especies de esponjas en esta foto. Esponja ramificada
Esponja ramificada La cesta de flores de Venus a una profundidad de 2572 metros
La cesta de flores de Venus a una profundidad de 2572 metros Esponja de barro
Esponja de barro La vida larga Monorhaphis chuni
La vida larga Monorhaphis chuni

.jpg)
.jpg)
.jpg)

Linneo identificó erróneamente las esponjas como plantas del orden de las algas. Durante mucho tiempo, las esponjas fueron asignadas a un subreino separado, Parazoa (que significa al lado de los animales). Ahora están clasificados como un filo parafilético a partir del cual han evolucionado los animales superiores.
Ctenóforos
Los ctenóforos (del griego, llevar un peine), comúnmente conocidos como medusas peine, son un filo que vive en aguas marinas en todo el mundo. Son los animales no coloniales más grandes que nadan con ayuda de cilios (pelos o peinetas). Las especies costeras deben ser lo suficientemente resistentes para resistir las olas y los remolinos de sedimentos, pero algunas especies oceánicas son tan frágiles y transparentes que es muy difícil capturarlas intactas para su estudio. En el pasado se pensaba que los ctenóforos tenían sólo una presencia modesta en el océano, pero ahora se sabe que a menudo son partes importantes e incluso dominantes de la biomasa planctónica.
El filo tiene alrededor de 150 especies conocidas con una amplia gama de formas corporales. Los tamaños varían desde unos pocos milímetros hasta 1,5 m (4 pies 11 pulgadas). Los cídípidos tienen forma de huevo con sus cilios dispuestos en ocho filas de peines radiales y despliegan tentáculos retráctiles para capturar presas. Los platicténidos bentónicos generalmente no tienen peine y son planos. Los beroides costeros tienen la boca abierta y carecen de tentáculos. La mayoría de los ctenóforos adultos se alimentan de larvas microscópicas, rotíferos y pequeños crustáceos, pero los beroides se alimentan de otros ctenóforos.
 Divulgación de luz a lo largo de las filas de peine de un tentáculo cidippid, izquierdo desplegado, retractado derecho
Divulgación de luz a lo largo de las filas de peine de un tentáculo cidippid, izquierdo desplegado, retractado derecho Los tentáculos profundos de citofora que se arrastran con tentilla (sustentáculos)
Los tentáculos profundos de citofora que se arrastran con tentilla (sustentáculos) Cydippid ctenophore en forma de huevo
Cydippid ctenophore en forma de huevo Grupo de pequeños gelatinas bentónicas que fluyen tentáculos y viven simbióticamente en un pez estrella.
Grupo de pequeños gelatinas bentónicas que fluyen tentáculos y viven simbióticamente en un pez estrella. Lobata con lóbulos gruesos pareados
Lobata con lóbulos gruesos pareados![The sea walnut has a transient anus which forms only when it needs to defecate.[223]](https://upload.wikimedia.org/wikipedia/commons/thumb/f/f8/Mnemiopsis_leidyi_2.jpg/255px-Mnemiopsis_leidyi_2.jpg) El nogal marino tiene un ano transitorio que se forma sólo cuando necesita defecar.
El nogal marino tiene un ano transitorio que se forma sólo cuando necesita defecar.



_on_Echniaster_luzonicus_(Seastar).jpg)

![The sea walnut has a transient anus which forms only when it needs to defecate.[223]](https://en.wikipedia.org/wiki/File:Mnemiopsis_leidyi_2.jpg)

Los primeros escritores combinaron ctenóforos con cnidarios. Los ctenóforos se parecen a los cnidarios en que dependen del flujo de agua a través de la cavidad corporal tanto para la digestión como para la respiración, así como en que tienen una red nerviosa descentralizada en lugar de un cerebro. También como los cnidarios, los cuerpos de los ctenóforos consisten en una masa de gelatina, con una capa de células en el exterior y otra que recubre la cavidad interna. En los ctenóforos, sin embargo, estas capas tienen dos células de profundidad, mientras que las de los cnidarios tienen solo una célula de profundidad. Mientras que los cnidarios exhiben simetría radial, los ctenóforos tienen dos canales anales que exhiben simetría biradial (simetría rotacional de media vuelta). La posición de los ctenóforos en el árbol genealógico evolutivo de los animales ha sido debatida durante mucho tiempo, y la opinión mayoritaria en la actualidad, basada en la filogenética molecular, es que los cnidarios y los bilaterianos están más estrechamente relacionados entre sí que cualquiera de ellos con los ctenóforos.
Placozoos
Los placozoos (del griego, animales planos) tienen la estructura más simple de todos los animales. Son una forma basal de organismo multicelular de vida libre (no parásito) que aún no tiene un nombre común. Viven en ambientes marinos y forman un filo que contiene hasta ahora solo tres especies descritas, de las cuales la primera, la clásica Trichoplax adhaerens, fue descubierta en 1883. Desde 2017 se han descubierto dos especies más, y los métodos genéticos indican que este filo tiene entre 100 y 200 especies más no descritas.



Trichoplax es un animal pequeño, aplanado, de aproximadamente un mm de ancho y generalmente alrededor de 25 µm de espesor. Al igual que las amebas a las que se parecen superficialmente, cambian continuamente su forma externa. Además, ocasionalmente se forman fases esféricas que pueden facilitar el movimiento. Trichoplax carece de tejidos y órganos. No hay simetría corporal manifiesta, por lo que no es posible distinguir la parte anterior de la posterior o la izquierda de la derecha. Está formado por unos miles de células de seis tipos en tres capas distintas. La capa exterior de células epiteliales simples tiene cilios que el animal utiliza para ayudarle a arrastrarse por el fondo marino. Trichoplax se alimenta engullendo y absorbiendo partículas de alimentos (principalmente microbios y detritos orgánicos) con su parte inferior.
Cnidarios marinos
_999_(30695685804).jpg)
Los cnidarios (del griego ortiga) se distinguen por la presencia de células urticantes, células especializadas que utilizan principalmente para capturar presas. Los cnidarios incluyen corales, anémonas de mar, medusas e hidrozoos. Forman un filo que contiene más de 10.000 especies de animales que se encuentran exclusivamente en ambientes acuáticos (principalmente marinos). Sus cuerpos están formados por mesoglea, una sustancia gelatinosa no viva, intercalada entre dos capas de epitelio que en su mayoría tienen un espesor de una célula. Tienen dos formas corporales básicas: medusas nadadoras y pólipos sésiles, ambos radialmente simétricos con bocas rodeadas de tentáculos que contienen cnidocitos. Ambas formas tienen un único orificio y cavidad corporal que se utilizan para la digestión y la respiración.
Se han encontrado cnidarios fósiles en rocas formadas hace unos hace unos 580 millones de años. Los fósiles de cnidarios que no forman estructuras mineralizadas son raros. Actualmente, los científicos piensan que los cnidarios, ctenóforos y bilaterianos están más estrechamente relacionados con las esponjas calcáreas que éstas con otras esponjas, y que los antozoos son las "tías" o "hermanas" de otros cnidarios, y el más estrechamente relacionado con los bilaterales.
Los cnidarios son los animales más simples en los que las células están organizadas en tejidos. La anémona de mar estrella se utiliza como organismo modelo en la investigación. Es fácil de cuidar en el laboratorio y se ha desarrollado un protocolo que puede producir una gran cantidad de embriones diariamente. Existe un grado notable de similitud en la conservación y complejidad de la secuencia genética entre la anémona de mar y los vertebrados. En particular, en la anémona también se encuentran genes implicados en la formación de la cabeza de los vertebrados.
 Los anemones de mar son comunes en las piscinas de marea.
Los anemones de mar son comunes en las piscinas de marea. Sus tentáculos pican y paralizan peces pequeños.
Sus tentáculos pican y paralizan peces pequeños. Cerca de pólipos en la superficie de un coral, agitando sus tentáculos.
Cerca de pólipos en la superficie de un coral, agitando sus tentáculos. Si una isla se hunde por debajo del mar, el crecimiento del coral puede mantenerse al día con el aumento del agua y formar un atolón.
Si una isla se hunde por debajo del mar, el crecimiento del coral puede mantenerse al día con el aumento del agua y formar un atolón.![The mantle of the red paper lantern jellyfish crumples and expands like a paper lantern.[239]](https://upload.wikimedia.org/wikipedia/commons/thumb/e/e5/Red-paper-lantern-jellyfish-Karen-Osborn-Smithsonian-Institution.png/321px-Red-paper-lantern-jellyfish-Karen-Osborn-Smithsonian-Institution.png) El manto de la linterna de papel rojo medusas y se expande como una linterna de papel.
El manto de la linterna de papel rojo medusas y se expande como una linterna de papel.




![The mantle of the red paper lantern jellyfish crumples and expands like a paper lantern.[239]](https://en.wikipedia.org/wiki/File:Red-paper-lantern-jellyfish-Karen-Osborn-Smithsonian-Institution.png)
 La guerra portuguesa es un sifonoforo colonial
La guerra portuguesa es un sifonoforo colonial Marrus orthocanna otro sifonóforo colonial, montado de dos tipos de zooides.
Marrus orthocanna otro sifonóforo colonial, montado de dos tipos de zooides.![Porpita porpita consists of a colony of hydroids[240]](https://upload.wikimedia.org/wikipedia/commons/thumb/2/23/Porpita_porpita.jpg/252px-Porpita_porpita.jpg) Porpita porpita consiste en una colonia de hidroides
Porpita porpita consiste en una colonia de hidroides![Lion's mane jellyfish, largest known jellyfish[241]](https://upload.wikimedia.org/wikipedia/commons/thumb/2/22/Largelionsmanejellyfish.jpg/191px-Largelionsmanejellyfish.jpg) Medusa de mane de león, medusa más conocida
Medusa de mane de león, medusa más conocida![Turritopsis dohrnii achieves biological immortality by transferring its cells back to childhood.[242][243]](https://upload.wikimedia.org/wikipedia/commons/b/b0/Turritopsis_dohrnii_%28cropped%29.jpg) Turritopsis dohrnii consigue la inmortalidad biológica transfiriendo sus células de vuelta a la infancia.
Turritopsis dohrnii consigue la inmortalidad biológica transfiriendo sus células de vuelta a la infancia.![The sea wasp is the most lethal jellyfish in the world.[244]](https://upload.wikimedia.org/wikipedia/commons/thumb/7/75/Chironex_fleckeri_%28sea_wasp%29.jpg/255px-Chironex_fleckeri_%28sea_wasp%29.jpg) La avispa marina es la medusa más letal del mundo.
La avispa marina es la medusa más letal del mundo.
.jpg)

![Porpita porpita consists of a colony of hydroids[240]](https://en.wikipedia.org/wiki/File:Porpita_porpita.jpg)
![Lion's mane jellyfish, largest known jellyfish[241]](https://en.wikipedia.org/wiki/File:Largelionsmanejellyfish.jpg)
![Turritopsis dohrnii achieves biological immortality by transferring its cells back to childhood.[242][243]](https://en.wikipedia.org/wiki/File:Turritopsis_dohrnii_(cropped).jpg)
![The sea wasp is the most lethal jellyfish in the world.[244]](https://en.wikipedia.org/wiki/File:Chironex_fleckeri_(sea_wasp).jpg)
Animales invertebrados bilaterales

Algunos de los primeros bilaterianos tenían forma de gusano, y el bilateriano original puede haber sido un gusano que habitaba en el fondo y tenía una sola abertura en el cuerpo. Un cuerpo bilateral puede conceptualizarse como un cilindro con un intestino que corre entre dos aberturas, la boca y el ano. Alrededor del intestino tiene una cavidad corporal interna, un celoma o pseudoceloma. Los animales con este plan corporal bilateralmente simétrico tienen un extremo de cabeza (anterior) y un extremo de cola (posterior), así como una espalda (dorsal) y un vientre (ventral); por lo tanto también tienen un lado izquierdo y un lado derecho.
Tener una parte delantera significa que esta parte del cuerpo encuentra estímulos, como la comida, favoreciendo la cefalización, el desarrollo de una cabeza con órganos sensoriales y una boca. El cuerpo se extiende hacia atrás desde la cabeza y muchos bilaterales tienen una combinación de músculos circulares que contraen el cuerpo, haciéndolo más largo, y un conjunto opuesto de músculos longitudinales, que acortan el cuerpo; estos permiten que los animales de cuerpo blando con esqueleto hidrostático se muevan mediante peristalsis. También tienen un intestino que se extiende a través del cuerpo básicamente cilíndrico desde la boca hasta el ano. Muchos filos bilaterales tienen larvas primarias que nadan con cilios y tienen un órgano apical que contiene células sensoriales. Sin embargo, existen excepciones a cada una de estas características; por ejemplo, los equinodermos adultos son radialmente simétricos (a diferencia de sus larvas) y ciertos gusanos parásitos tienen estructuras corporales extremadamente simplificadas.
.jpg)
| ← bilaterios |
| |||||||||||||||||||||
Protóstomos
Los protóstomos (del griego, primera boca) son un superfilo de animales. Es un clado hermano de los deuteróstomos (del griego segunda boca), con el que forma el clado Nephrozoa. Los protóstomos se distinguen de los deuteróstomos por la forma en que se desarrollan sus embriones. En los protóstomos la primera abertura que se desarrolla se convierte en la boca, mientras que en los deuteróstomos se convierte en el ano.
| ← Protostomes |
| ||||||||||||||||||||||||||||||||||||||||||
| (extante) |


Gusanos marinos

Los gusanos (en inglés antiguo, serpientes) forman varios filos. Los diferentes grupos de gusanos marinos están relacionados sólo de forma lejana, por lo que se encuentran en varios filos diferentes, como Annelida (gusanos segmentados), Chaetognatha (gusanos flecha), Phoronida (gusanos de herradura) y Hemichordata. Todos los gusanos, excepto los Hemichordata, son protóstomos. Los hemicordados son deuteróstomos y se analizan en su propia sección a continuación.
El diseño corporal típico de un gusano implica cuerpos largos y cilíndricos en forma de tubo y sin extremidades. Los gusanos marinos varían en tamaño, desde microscópicos hasta más de 1 metro (3,3 pies) de longitud para algunos gusanos poliquetos marinos (gusanos de cerdas) y hasta 58 metros (190 pies) para el gusano nemertino marino (gusano de cordón). Algunos gusanos marinos ocupan una pequeña variedad de nichos parásitos y viven dentro de los cuerpos de otros animales, mientras que otros viven más libremente en el ambiente marino o excavando bajo tierra. Muchos de estos gusanos tienen tentáculos especializados que se utilizan para intercambiar oxígeno y dióxido de carbono y también pueden usarse para la reproducción. Algunos gusanos marinos son gusanos tubulares, como el gusano tubular gigante que vive en aguas cercanas a volcanes submarinos y puede soportar temperaturas de hasta 90 grados centígrados. Los platelmintos (gusanos planos) forman otro filo de gusanos que incluye una clase de tenias parásitas. La tenia marina Polygonoporus giganticus, que se encuentra en el intestino de los cachalotes, puede crecer hasta más de 30 m (100 pies).
Los nematodos (gusanos redondos) constituyen otro filo de gusanos con sistemas digestivos tubulares y una abertura en ambos extremos. Se han descrito más de 25.000 especies de nematodos, de las cuales más de la mitad son parásitos. Se estima que otro millón sigue sin ser descrito. Son omnipresentes en ambientes marinos, de agua dulce y terrestres, donde a menudo superan en número a otros animales tanto en número de individuos como de especies. Se encuentran en todas partes de la litosfera de la Tierra, desde la cima de las montañas hasta el fondo de las fosas oceánicas. Según su conteo, representan el 90% de todos los animales del fondo del océano. Su dominancia numérica, que a menudo supera el millón de individuos por metro cuadrado y representa alrededor del 80% de todos los animales individuales de la Tierra, su diversidad de ciclos de vida y su presencia en varios niveles tróficos apuntan a un papel importante en muchos ecosistemas.
 Los gusanos gigantes de tubo se agrupan alrededor de los respiraderos hidrotermales.
Los gusanos gigantes de tubo se agrupan alrededor de los respiraderos hidrotermales. Los nematodos son pseudocoelomatos ubicuos que pueden parásitos plantas y animales marinos.
Los nematodos son pseudocoelomatos ubicuos que pueden parásitos plantas y animales marinos. Los gusanos de sangre se encuentran típicamente en el fondo de aguas marinas poco profundas.
Los gusanos de sangre se encuentran típicamente en el fondo de aguas marinas poco profundas.



Moluscos marinos




Los moluscos (del latín blando) forman un filo con alrededor de 85.000 especies reconocidas. Son el filo marino más grande en términos de número de especies y contienen aproximadamente el 23% de todos los organismos marinos nombrados. Los moluscos tienen formas más variadas que otros filos de invertebrados. Son muy diversos, no sólo en tamaño y estructura anatómica, sino también en comportamiento y hábitat.
.jpg)
El filo de los moluscos se divide en 9 o 10 clases taxonómicas. Estas clases incluyen gasterópodos, bivalvos y cefalópodos, así como otras clases menos conocidas pero distintivas. Los gasterópodos con caparazón protector se denominan caracoles, mientras que los gasterópodos sin caparazón protector se denominan babosas. Los gasterópodos son, con diferencia, los moluscos más numerosos en términos de especies. Los bivalvos incluyen almejas, ostras, berberechos, mejillones, vieiras y muchas otras familias. Hay alrededor de 8.000 especies de bivalvos marinos (incluidas especies de aguas salobres y de estuarios). Se ha informado que una almeja quahog de aguas profundas vivió 507 años, lo que la convierte en la vida más larga registrada de todos los animales, aparte de los animales coloniales o animales casi coloniales como las esponjas.
- Gastropods and bivalves
 Los gastropods marinos son caracoles de mar o langostas de mar. Este nudibranch es un bazo de mar.
Los gastropods marinos son caracoles de mar o langostas de mar. Este nudibranch es un bazo de mar. El caracol del mar Syrinx aruanus tiene una cáscara de hasta 91 cm de largo, el mayor de cualquier gastropo viviente.
El caracol del mar Syrinx aruanus tiene una cáscara de hasta 91 cm de largo, el mayor de cualquier gastropo viviente. Los moluscos suelen tener ojos. Bordeando el borde del manto de un scallop, un bivalve mollusc, puede ser más de 100 ojos simples.
Los moluscos suelen tener ojos. Bordeando el borde del manto de un scallop, un bivalve mollusc, puede ser más de 100 ojos simples. Mejillón común, otro bivalvo
Mejillón común, otro bivalvo




Los cefalópodos incluyen pulpos, calamares y sepias. Se han identificado unas 800 especies vivas de cefalópodos marinos y se han descrito unos 11.000 taxones extintos. Se encuentran en todos los océanos, pero no existen cefalópodos totalmente de agua dulce.
- Cefalópodos
![The nautilus is a living fossil little changed since it evolved 500 million years ago as one of the first cephalopods.[274][275][276]](https://upload.wikimedia.org/wikipedia/commons/thumb/d/d5/Nautilus_Palau.JPG/300px-Nautilus_Palau.JPG) El nautilus es un fósil vivo poco cambiado desde que evolucionó hace 500 millones de años como uno de los primeros cefalopodos.
El nautilus es un fósil vivo poco cambiado desde que evolucionó hace 500 millones de años como uno de los primeros cefalopodos. Reconstrucción de un ammonito, un cefalopodo temprano muy exitoso que apareció 400 mia.
Reconstrucción de un ammonito, un cefalopodo temprano muy exitoso que apareció 400 mia. Los cefalopodos, como este pez cuttle, usan su cavidad de manto para propulsión jet.
Los cefalopodos, como este pez cuttle, usan su cavidad de manto para propulsión jet.![Colossal squid, the largest of all invertebrates[277]](https://upload.wikimedia.org/wikipedia/commons/thumb/3/37/Calmarcolossal.jpg/157px-Calmarcolossal.jpg) Colosal calamar, el más grande de todos los invertebrados
Colosal calamar, el más grande de todos los invertebrados
![The nautilus is a living fossil little changed since it evolved 500 million years ago as one of the first cephalopods.[274][275][276]](https://en.wikipedia.org/wiki/File:Nautilus_Palau.JPG)


![Colossal squid, the largest of all invertebrates[277]](https://en.wikipedia.org/wiki/File:Calmarcolossal.jpg)
Los moluscos tienen formas tan diversas que muchos libros de texto basan sus descripciones de la anatomía de los moluscos en un molusco ancestral generalizado o hipotético. Este molusco generalizado no está segmentado y es bilateralmente simétrico con una parte inferior que consta de un solo pie musculoso. Más allá de eso, tiene otras tres características clave. En primer lugar, tiene un manto muscular llamado manto que cubre sus vísceras y contiene una importante cavidad que utiliza para respirar y excretar. Una concha secretada por el manto cubre la superficie superior. En segundo lugar (aparte de los bivalvos) tiene una lengua áspera llamada rádula que utiliza para alimentarse. En tercer lugar, tiene un sistema nervioso que incluye un sistema digestivo complejo que utiliza pelos microscópicos impulsados por músculos llamados cilios para exudar moco. El molusco generalizado tiene dos cordones nerviosos pares (tres en los bivalvos). El cerebro, en las especies que lo tienen, rodea el esófago. La mayoría de los moluscos tienen ojos y todos tienen sensores que detectan sustancias químicas, vibraciones y tacto.
Existe buena evidencia de la aparición de gasterópodos, cefalópodos y bivalvos marinos en el período Cámbrico hace 538,8 a 485,4 millones de años.
Artrópodos marinos


Los artrópodos (del griego, pies articulados) tienen un exoesqueleto (esqueleto externo), un cuerpo segmentado y apéndices articulados (apéndices pareados). Forman un filo que incluye insectos, arácnidos, miriápodos y crustáceos. Los artrópodos se caracterizan por sus extremidades articuladas y una cutícula hecha de quitina, a menudo mineralizada con carbonato de calcio. El plan corporal de los artrópodos consta de segmentos, cada uno con un par de apéndices. La cutícula rígida inhibe el crecimiento, por lo que los artrópodos la reemplazan periódicamente mediante mudas. Su versatilidad les ha permitido convertirse en los miembros más ricos en especies de todos los gremios ecológicos en la mayoría de los entornos.
La ascendencia evolutiva de los artrópodos se remonta al período Cámbrico y generalmente se considera monofilética. Sin embargo, recientemente se han debatido las relaciones basales de los artrópodos con filos extintos como los lobopodios.
| Panarthropoda |
| ||||||||||||


- Fósiles y fósiles vivos
![Fossil trilobite. Trilobites first appeared about 521 Ma. They were highly successful and were found everywhere in the ocean for 270 Ma.[285]](https://upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Cheirurus_ingricus.png/301px-Cheirurus_ingricus.png) Fossil trilobite. Trilobites apareció por primera vez alrededor de 521 Ma. Fueron muy exitosos y fueron encontrados en todas partes en el océano para 270 Ma.
Fossil trilobite. Trilobites apareció por primera vez alrededor de 521 Ma. Fueron muy exitosos y fueron encontrados en todas partes en el océano para 270 Ma. El Anomalocaris ("barrón anormal") fue uno de los primeros depredadores de ápice y apareció por primera vez alrededor de 515 Ma.
El Anomalocaris ("barrón anormal") fue uno de los primeros depredadores de ápice y apareció por primera vez alrededor de 515 Ma.![The largest known arthropod, the sea scorpion Jaekelopterus rhenaniae, has been found in estuarine strata from about 390 Ma. It was up to 2.5 m (8.2 ft) long.[286][287]](https://upload.wikimedia.org/wikipedia/commons/thumb/a/af/Jaekelopterus_rhenaniae_reconstruction.jpg/331px-Jaekelopterus_rhenaniae_reconstruction.jpg) El artrópodo más grande conocido, el escorpión del mar Jaekelopterus rhenaniae, se ha encontrado en estuarine strata de aproximadamente 390 Ma. Era de hasta 2,5 m (8.2 pies) de largo.
El artrópodo más grande conocido, el escorpión del mar Jaekelopterus rhenaniae, se ha encontrado en estuarine strata de aproximadamente 390 Ma. Era de hasta 2,5 m (8.2 pies) de largo.![Xiphosurans, the group including modern Horseshoe crabs appeared around 480 Ma.[288]](https://upload.wikimedia.org/wikipedia/commons/thumb/d/d4/Limulus_%28cropped%29.jpg/257px-Limulus_%28cropped%29.jpg) Xiphosurans, el grupo incluyendo modernos cangrejos Horseshoe apareció alrededor de 480 Ma.
Xiphosurans, el grupo incluyendo modernos cangrejos Horseshoe apareció alrededor de 480 Ma.
![Fossil trilobite. Trilobites first appeared about 521 Ma. They were highly successful and were found everywhere in the ocean for 270 Ma.[285]](https://en.wikipedia.org/wiki/File:Cheirurus_ingricus.png)

![The largest known arthropod, the sea scorpion Jaekelopterus rhenaniae, has been found in estuarine strata from about 390 Ma. It was up to 2.5 m (8.2 ft) long.[286][287]](https://en.wikipedia.org/wiki/File:Jaekelopterus_rhenaniae_reconstruction.jpg)
![Xiphosurans, the group including modern Horseshoe crabs appeared around 480 Ma.[288]](https://en.wikipedia.org/wiki/File:Limulus_(cropped).jpg)
Los artrópodos marinos existentes varían en tamaño desde el crustáceo microscópico Stygotantulus hasta el cangrejo araña japonés. Artrópodos' la cavidad interna primaria es un hemocele, que alberga sus órganos internos y a través del cual circula su hemolinfa, un análogo de la sangre; Tienen sistemas circulatorios abiertos. Al igual que su exterior, los órganos internos de los artrópodos generalmente están formados por segmentos repetidos. Su sistema nervioso tiene forma de escalera, con pares de cordones nerviosos ventrales que recorren todos los segmentos y forman pares de ganglios en cada segmento. Sus cabezas se forman mediante la fusión de un número variable de segmentos, y sus cerebros se forman mediante la fusión de los ganglios de estos segmentos y rodean el esófago. Los sistemas respiratorio y excretor de los artrópodos varían, dependiendo tanto de su entorno como del subfilo al que pertenecen.
- Modernos crustáceos
 Muchos crustáceos son muy pequeños, como este pequeño anfipodo, y constituyen una parte significativa del zooplancton del océano.
Muchos crustáceos son muy pequeños, como este pequeño anfipodo, y constituyen una parte significativa del zooplancton del océano.![The Japanese spider crab has the longest leg span of any arthropod, reaching 5.5 metres (18 ft) from claw to claw.[289]](https://upload.wikimedia.org/wikipedia/commons/thumb/a/a7/Macrocheira_kaempferi.jpg/314px-Macrocheira_kaempferi.jpg) El cangrejo de araña japonesa tiene el más largo lapso de pierna de cualquier artrópodo, alcanzando 5,5 metros (18 pies) de garra a garra.
El cangrejo de araña japonesa tiene el más largo lapso de pierna de cualquier artrópodo, alcanzando 5,5 metros (18 pies) de garra a garra.![The Tasmanian giant crab is long-lived and slow-growing, making it vulnerable to overfishing.[290]](https://upload.wikimedia.org/wikipedia/commons/thumb/2/2c/J_J_Wild_Pseudocarcinus_cropped.jpg/330px-J_J_Wild_Pseudocarcinus_cropped.jpg) El cangrejo gigante de Tasmania es de larga vida y lento crecimiento, lo que lo hace vulnerable a la sobrepesca.
El cangrejo gigante de Tasmania es de larga vida y lento crecimiento, lo que lo hace vulnerable a la sobrepesca.![Mantis shrimp have the most advanced eyes in the animal kingdom,[291] and smash prey by swinging their club-like raptorial claws.[292]](https://upload.wikimedia.org/wikipedia/commons/thumb/0/03/Odontodactylus_scyllarus_2.png/298px-Odontodactylus_scyllarus_2.png) Los camarones Mantis tienen los ojos más avanzados en el reino animal, y rompen presas colgando sus garras raptoriales tipo club.
Los camarones Mantis tienen los ojos más avanzados en el reino animal, y rompen presas colgando sus garras raptoriales tipo club.

![The Japanese spider crab has the longest leg span of any arthropod, reaching 5.5 metres (18 ft) from claw to claw.[289]](https://en.wikipedia.org/wiki/File:Macrocheira_kaempferi.jpg)
![The Tasmanian giant crab is long-lived and slow-growing, making it vulnerable to overfishing.[290]](https://en.wikipedia.org/wiki/File:J_J_Wild_Pseudocarcinus_cropped.jpg)
![Mantis shrimp have the most advanced eyes in the animal kingdom,[291] and smash prey by swinging their club-like raptorial claws.[292]](https://en.wikipedia.org/wiki/File:Odontodactylus_scyllarus_2.png)
La visión de los artrópodos se basa en varias combinaciones de ojos compuestos y ocelos con fosas pigmentarias: en la mayoría de las especies, los ocelos sólo pueden detectar la dirección de donde proviene la luz, y los ojos compuestos son la principal fuente de información. Los artrópodos también tienen una amplia gama de sensores químicos y mecánicos, basados principalmente en modificaciones de las numerosas setas (cerdas) que se proyectan a través de sus cutículas. Los métodos de reproducción de los artrópodos son diversos: las especies terrestres utilizan alguna forma de fertilización interna, mientras que las especies marinas ponen huevos mediante fertilización interna o externa. Las crías de artrópodos varían desde adultos en miniatura hasta larvas que carecen de extremidades articuladas y eventualmente sufren una metamorfosis total para producir la forma adulta.
Deuteróstomos
En los deuteróstomos, la primera abertura que se desarrolla en el embrión en crecimiento se convierte en el ano, mientras que en los protóstomos se convierte en la boca. Los deuterostomos forman un superfilo de animales y son el clado hermano de los protostomos. Alguna vez se consideró que los primeros deuteróstomos conocidos son fósiles de Saccorhytus de hace unos 540 millones de años. Sin embargo, otro estudio consideró que es más probable que Saccorhytus sea un ecdisozoo.
| ← deuterostomía |
| ||||||||||||||||||||||||
| (extante) |

Equinodermos

Los equinodermos (del griego, piel espinosa) son un filo que contiene únicamente invertebrados marinos. El filo contiene alrededor de 7000 especies vivas, lo que lo convierte en el segundo grupo más grande de deuteróstomos, después de los cordados.
Los equinodermos adultos son reconocibles por su simetría radial (generalmente de cinco puntos) e incluyen estrellas de mar, erizos de mar, dólares de arena y pepinos de mar, así como lirios de mar. Los equinodermos se encuentran en todas las profundidades del océano, desde la zona intermareal hasta la zona abisal. Son únicos entre los animales por tener simetría bilateral en la etapa larvaria, pero simetría quíntuple (pentamerismo, un tipo especial de simetría radial) en la edad adulta.
Los equinodermos son importantes tanto biológica como geológicamente. Biológicamente, hay pocos otros grupos tan abundantes en el desierto biótico de las profundidades del mar, así como en los océanos menos profundos. La mayoría de los equinodermos son capaces de regenerar tejidos, órganos, extremidades y reproducirse asexualmente; en algunos casos, pueden sufrir una regeneración completa a partir de una sola extremidad. Geológicamente, el valor de los equinodermos reside en sus esqueletos osificados, que contribuyen en gran medida a muchas formaciones de piedra caliza y pueden proporcionar pistas valiosas sobre el entorno geológico. Fueron las especies más utilizadas en la investigación regenerativa en los siglos XIX y XX.
 Echinoderm literalmente significa "spiny skin", como ilustra este erizo de melón de agua.
Echinoderm literalmente significa "spiny skin", como ilustra este erizo de melón de agua.![The ochre sea star was the first keystone predator to be studied. They limit mussels which can overwhelm intertidal communities.[298]](https://upload.wikimedia.org/wikipedia/commons/thumb/8/89/Ochre_sea_star.jpg/360px-Ochre_sea_star.jpg) La estrella del mar ocre fue el primer depredador de piedra clave para ser estudiado. Limitan los mejillones que pueden abrumar a las comunidades intermareales.
La estrella del mar ocre fue el primer depredador de piedra clave para ser estudiado. Limitan los mejillones que pueden abrumar a las comunidades intermareales. Lirios de mar coloridos en aguas poco profundas
Lirios de mar coloridos en aguas poco profundas

![The ochre sea star was the first keystone predator to be studied. They limit mussels which can overwhelm intertidal communities.[298]](https://en.wikipedia.org/wiki/File:Ochre_sea_star.jpg)

 Los pepinos de mar filtro alimentan en plancton y sólidos suspendidos.
Los pepinos de mar filtro alimentan en plancton y sólidos suspendidos. El cerdo de mar, un pepino de agua profunda, es el único equinodermo que utiliza la locomoción legged.
El cerdo de mar, un pepino de agua profunda, es el único equinodermo que utiliza la locomoción legged. Un pepino de mar de natación benéopelagico y bioluminiscente, 3200 metros de profundidad
Un pepino de mar de natación benéopelagico y bioluminiscente, 3200 metros de profundidad

.jpg)

Algunos científicos sostienen que la radiación de los equinodermos fue responsable de la Revolución Marina Mesozoica. Aparte del difícil de clasificar Arkarua (un animal precámbrico con simetría radial pentámera similar a un equinodermo), los primeros miembros definitivos del filo aparecieron cerca del comienzo del Cámbrico.
Hemicordados


Los hemicordados forman un filo hermano de los equinodermos. Son organismos solitarios con forma de gusano que los humanos rara vez ven debido a su estilo de vida. Incluyen dos grupos principales, los gusanos belloteros y los pterobranquios. Los pterobranquios forman una clase que contiene alrededor de 30 especies de pequeños animales con forma de gusano que viven en tubos secretados en el fondo del océano. Los gusanos belloteros forman una clase que contiene alrededor de 111 especies y generalmente viven en madrigueras en forma de U en el fondo marino, desde la costa hasta una profundidad de 3.000 metros. Los gusanos yacen allí con la probóscide sobresaliendo de una abertura de la madriguera, subsistiendo como alimentos de depósito o de suspensión. Se supone que los antepasados de los gusanos belloteros solían vivir en tubos como sus parientes, los pterobranchia, pero finalmente comenzaron a vivir una existencia más segura y protegida en madrigueras de sedimentos. Algunos de estos gusanos pueden llegar a ser muy largos; una especie en particular puede alcanzar una longitud de 2,5 metros (8 pies 2 pulgadas), aunque la mayoría de los gusanos belloteros son mucho más pequeños.
Los gusanos bellota son más especializados y avanzados que otros organismos parecidos a gusanos. Tienen un sistema circulatorio con un corazón que también funciona como riñón. Los gusanos belloteros tienen estructuras parecidas a branquias que utilizan para respirar, similares a las branquias de los peces. Por lo tanto, a veces se dice que los gusanos belloteros son un vínculo entre los invertebrados clásicos y los vertebrados. Los gusanos belloteros forman continuamente nuevas hendiduras branquiales a medida que crecen en tamaño, y algunos individuos mayores tienen más de cien en cada lado. Cada hendidura consta de una cámara branquial que se abre a la faringe a través de una hendidura en forma de U. Los cilios empujan el agua a través de las rendijas, manteniendo un flujo constante, como en los peces. Algunos gusanos belloteros también tienen una cola postanal que puede ser homóloga a la cola postanal de los vertebrados.
El plan corporal de tres secciones del gusano bellotero ya no está presente en los vertebrados, excepto en la anatomía del tubo neural frontal, que luego se desarrolló hasta convertirse en un cerebro dividido en tres partes. Esto significa que parte de la anatomía original de los primeros ancestros cordados todavía está presente en los vertebrados, aunque no siempre sea visible. Una teoría es que el cuerpo de tres partes se originó a partir de un ancestro común temprano de los deuteróstomos, y tal vez incluso de un ancestro común bilateral tanto de los deuteróstomos como de los protóstomos. Los estudios han demostrado que la expresión genética en el embrión comparte tres de los mismos centros de señalización que dan forma al cerebro de todos los vertebrados, pero en lugar de participar en la formación de su sistema neuronal, controlan el desarrollo de las diferentes regiones del cuerpo.
Cordadas marinas
(feminine)
El filo de los cordados tiene tres subfilos, uno de los cuales son los vertebrados (ver más abajo). Los otros dos subfilos son invertebrados marinos: los tunicados (salpas y ascidias) y los cefalocordados (como las lancetas). Los cordados invertebrados son parientes cercanos de los vertebrados. En particular, se ha debatido sobre qué tan estrechamente algunas especies marinas extintas, como Pikaiidae, Palaeospondylus, Zhongxiniscus y Vetulicolia, podrían relacionarse ancestralmente con los vertebrados.
- Los acordes invertebrados son parientes cercanos de vertebrados
![The lancelet, a small translucent fish-like cephalochordate, is one of the closest living invertebrate relative of the vertebrates.[307][308]](https://upload.wikimedia.org/wikipedia/commons/thumb/5/54/Amphioxus.png/332px-Amphioxus.png) El lancelet, un pequeño cefalochordate de peces translúcidos, es uno de los parientes invertebrados vivos más cercanos de los vertebrados.
El lancelet, un pequeño cefalochordate de peces translúcidos, es uno de los parientes invertebrados vivos más cercanos de los vertebrados.![Tunicates, like these fluorescent-colored sea squirts, may provide clues to vertebrate and therefore human ancestry.[309]](https://upload.wikimedia.org/wikipedia/commons/thumb/8/8c/Ascidian_%28Rhopalaea_Crassa%29_%284_cm%29.png/280px-Ascidian_%28Rhopalaea_Crassa%29_%284_cm%29.png) Los tunicatos, como estos chorros de mar de color fluorescente, pueden proporcionar pistas a los vertebrados y por lo tanto a la ascendencia humana.
Los tunicatos, como estos chorros de mar de color fluorescente, pueden proporcionar pistas a los vertebrados y por lo tanto a la ascendencia humana. Los pirosomas son tunicatos bioluminescentes de planta libre formados por cientos de individuos.
Los pirosomas son tunicatos bioluminescentes de planta libre formados por cientos de individuos. Cadena de salp
Cadena de salp
![The lancelet, a small translucent fish-like cephalochordate, is one of the closest living invertebrate relative of the vertebrates.[307][308]](https://en.wikipedia.org/wiki/File:Amphioxus.png)
![Tunicates, like these fluorescent-colored sea squirts, may provide clues to vertebrate and therefore human ancestry.[309]](https://en.wikipedia.org/wiki/File:Ascidian_(Rhopalaea_Crassa)_(4_cm).png)

.jpg)



Animales vertebrados


Los vertebrados (del latín articulaciones de la columna) son un subfilo de los cordados. Son cordados que tienen una columna vertebral (columna vertebral). La columna vertebral proporciona la estructura de soporte central para un esqueleto interno que da forma, soporte y protección al cuerpo y puede proporcionar un medio para anclar aletas o extremidades al cuerpo. La columna vertebral también sirve para albergar y proteger la médula espinal que se encuentra dentro de la columna vertebral.
Los vertebrados marinos se pueden dividir en peces marinos y tetrápodos marinos.
Pez marino
Los peces normalmente respiran extrayendo oxígeno del agua a través de branquias y tienen una piel protegida por escamas y mucosas. Utilizan aletas para impulsarse y estabilizarse en el agua y, por lo general, tienen un corazón de dos cámaras y ojos bien adaptados para ver bajo el agua, así como otros sistemas sensoriales. Hasta 2017 se han descrito más de 33.000 especies de peces, de las cuales unas 20.000 son peces marinos.
| ← vertebrados |
| |||||||||||||||||||||||||||
| (extante) |
Pez sin mandíbula

Los primeros peces no tenían mandíbulas. La mayoría se extinguió cuando fueron superados por los peces con mandíbulas (abajo), pero dos grupos sobrevivieron: los mixinos y las lampreas. Los mixinos forman una clase de unas 20 especies de peces marinos que producen limo y tienen forma de anguila. Son los únicos animales vivos conocidos que tienen cráneo pero no columna vertebral. Las lampreas forman una superclase que contiene 38 especies conocidas de peces sin mandíbula. La lamprea adulta se caracteriza por una boca chupadora dentada en forma de embudo. Aunque son bien conocidas por perforar la carne de otros peces para chuparles la sangre, sólo 18 especies de lampreas son realmente parásitas. Juntos, los mixinos y las lampreas son el grupo hermano de los vertebrados. Los mixinos vivos siguen siendo similares a los mixinos de hace unos 300 millones de años. Las lampreas son un linaje muy antiguo de vertebrados, aunque su relación exacta con los mixinos y los vertebrados con mandíbulas sigue siendo motivo de controversia. El análisis molecular desde 1992 ha sugerido que los mixinos están más estrechamente relacionados con las lampreas, y también lo están con los vertebrados en un sentido monofilético. Otros los consideran un grupo hermano de vertebrados en el taxón común de craniata.
El monstruo de Tully es un género extinto de bilaterales de cuerpo blando que vivió en estuarios tropicales hace unos 300 millones de años. Desde 2016 existe controversia sobre si este animal era un vertebrado o un invertebrado. En 2020, los investigadores encontraron "evidencias sólidas" que el monstruo de Tully era un vertebrado y era un pez sin mandíbula del linaje de la lamprea, mientras que en 2023 otros investigadores descubrieron que los escaneos de fósiles en 3D no respaldaban esas conclusiones.
 Los peces hagfish son los únicos animales vivos conocidos con un cráneo pero sin columna vertebral.
Los peces hagfish son los únicos animales vivos conocidos con un cráneo pero sin columna vertebral. Los lampreys son a menudo parasitarios y tienen una boca de chupadora de embudo dentada.
Los lampreys son a menudo parasitarios y tienen una boca de chupadora de embudo dentada. El extinto Pteraspidomorphi, ancestral a los vertebrados jawed
El extinto Pteraspidomorphi, ancestral a los vertebrados jawed



Pteraspidomorphi es una clase extinta de peces sin mandíbulas, ancestrales de los vertebrados con mandíbulas. Las pocas características que comparten con estos últimos se consideran ahora primitivas para todos los vertebrados.
Al inicio del Devónico, comenzaron a aparecer peces con una profunda remodelación del cráneo de los vertebrados que resultó en una mandíbula. Todas las mandíbulas de los vertebrados, incluida la mandíbula humana, han evolucionado a partir de estas primeras mandíbulas de peces. La aparición de la mandíbula de los primeros vertebrados se ha descrito como "quizás el paso evolutivo más profundo y radical en la historia de los vertebrados". Las mandíbulas permiten capturar, sostener y masticar presas. Los peces sin mandíbulas tuvieron más dificultades para sobrevivir que los peces con mandíbulas, y la mayoría de los peces sin mandíbulas se extinguieron durante el período Triásico.
Pez cartilaginoso
Los peces con mandíbulas se dividen en dos grupos principales: peces con esqueleto interno óseo y peces con esqueleto interno cartilaginoso. Los peces cartilaginosos, como los tiburones y las rayas, tienen mandíbulas y esqueletos hechos de cartílago en lugar de hueso. Megalodon es una especie extinta de tiburón que vivió entre 28 y 1,5 millones de años. Puede que se pareciera mucho a una versión robusta del gran tiburón blanco, pero era mucho más grande, con longitudes estimadas que alcanzaban los 20,3 metros (67 pies). Se encuentra en todos los océanos y fue uno de los depredadores más grandes y poderosos de la historia de los vertebrados, y probablemente tuvo un profundo impacto en la vida marina. El tiburón de Groenlandia tiene la esperanza de vida más larga conocida de todos los vertebrados, unos 400 años. Algunos tiburones, como el gran blanco, son parcialmente de sangre caliente y dan a luz vivos. La mantarraya, la raya más grande del mundo, ha sido objeto de pesca y ahora es vulnerable.
- Peces cartilaginosos
 Los peces cartilaginosos pueden haber evolucionado de tiburones espinosos.
Los peces cartilaginosos pueden haber evolucionado de tiburones espinosos. Stingray
Stingray Manta Ray, el rayo más grande
Manta Ray, el rayo más grande![Sawfish, rays with long rostrums resembling a saw. All species are now endangered.[332]](https://upload.wikimedia.org/wikipedia/commons/thumb/4/48/Pristis_clavata_2.jpg/407px-Pristis_clavata_2.jpg) Sawfish, rayos con largas tribunas que parecen una sierra. Todas las especies están ahora en peligro.
Sawfish, rayos con largas tribunas que parecen una sierra. Todas las especies están ahora en peligro.



![Sawfish, rays with long rostrums resembling a saw. All species are now endangered.[332]](https://en.wikipedia.org/wiki/File:Pristis_clavata_2.jpg)
 El extinto megalodón se parecía a un gran tiburón blanco gigante.
El extinto megalodón se parecía a un gran tiburón blanco gigante. El tiburón de Groenlandia vive más tiempo que cualquier otro vertebrado.
El tiburón de Groenlandia vive más tiempo que cualquier otro vertebrado. El pez extante más grande, el tiburón ballena, es ahora una especie vulnerable.
El pez extante más grande, el tiburón ballena, es ahora una especie vulnerable.


.jpg)
Pez óseo


.jpg)
Los peces óseos tienen mandíbulas y esqueletos hechos de hueso en lugar de cartílago. Los peces óseos también tienen placas óseas duras llamadas opérculo que les ayudan a respirar y proteger sus branquias y, a menudo, poseen una vejiga natatoria que utilizan para controlar mejor su flotabilidad. Los peces óseos se pueden dividir en aquellos con aletas lobuladas y aquellos con aletas radiales. Las fechas aproximadas en el árbol filogenético son de Near et al., 2012 y Zhu et al., 2009.
| ← pez bony |
| |||||||||||||||||||||||||||||||||
| (extante) |
Las aletas lobuladas tienen la forma de lóbulos carnosos sostenidos por tallos óseos que se extienden desde el cuerpo. Guiyu oneiros, el pez óseo más antiguo conocido, vivió durante el Silúrico Superior, hace 419 millones de años. Tiene la combinación de características de aletas radiadas y lobuladas, aunque el análisis de la totalidad de sus características lo ubica más cerca de los peces con aletas lobuladas. Las aletas lobuladas evolucionaron hasta convertirse en las patas de los primeros vertebrados terrestres tetrápodos, por lo que, por extensión, uno de los primeros antepasados de los humanos fue un pez con aletas lobuladas. Aparte de los celacantos y los peces pulmonados, los peces con aletas lobuladas están actualmente extintos.
Los peces óseos restantes tienen aletas radiadas. Están hechos de redes de piel sostenidas por espinas (rayos) óseas o córneas que pueden erigirse para controlar la rigidez de las aletas.
- La principal característica distintiva de los condrosteanos (esturgeon, paddlefish, bichir y reedfish) es la naturaleza cartilaginosa de sus esqueletos. Se cree que los ancestros de los condrosteanos son peces bolos, pero la característica de un esqueleto osificado se perdió en desarrollo evolutivo posterior, lo que dio lugar a un relámpago del marco.
- Neopterygians (de griego para nuevas aletas) apareció en algún momento en el Permian tardío, antes de los dinosaurios. Eran un grupo de peces muy exitoso, porque podían moverse más rápidamente que sus antepasados. Sus escamas y esqueletos comenzaron a iluminarse durante su evolución, y sus mandíbulas se volvieron más poderosas y eficientes.
Teleosteos

Aproximadamente el 96% de todas las especies de peces modernas son teleósteos, de los cuales alrededor de 14.000 son especies marinas. Los teleósteos se pueden distinguir de otros peces óseos por la posesión de una cola homocercal, una cola donde la mitad superior refleja la mitad inferior. Otra diferencia radica en los huesos de su mandíbula: los teleósteos tienen modificaciones en la musculatura de la mandíbula que les permite sobresalir. Esto les permite agarrar presas y llevárselas a la boca. En general, los teleósteos tienden a ser más rápidos y flexibles que los peces óseos más basales. Su estructura esquelética ha evolucionado hacia una mayor ligereza. Si bien los huesos de los teleósteos están bien calcificados, están construidos a partir de un andamiaje de puntales, en lugar de los densos huesos esponjosos de los peces holosteos.
Los teleósteos se encuentran en casi todos los hábitats marinos. Tienen una enorme diversidad y varían en tamaño desde gobios adultos de 8 mm de largo hasta peces luna que pesan más de 2000 kg. Las siguientes imágenes muestran algo de la diversidad en la forma y el color de los teleósteos marinos modernos...
 Sailfish
Sailfish Mahi-mahi
Mahi-mahi Eel
Eel Seahorse
Seahorse



 Ocean sunfish
Ocean sunfish Anglerfish
Anglerfish Pufferfish
Pufferfish Peces de gatillo payaso
Peces de gatillo payaso Dragonet de mandarina
Dragonet de mandarina





Casi la mitad de todas las especies de vertebrados existentes son teleósteos.
Tetrápodos marinos

Un tetrápodo (del griego, cuatro pies) es un vertebrado con extremidades (pies). Los tetrápodos evolucionaron a partir de antiguos peces con aletas lobuladas hace unos 400 millones de años durante el período Devónico, cuando sus primeros ancestros emergieron del mar y se adaptaron a vivir en la tierra. Este cambio de un plan corporal para respirar y navegar en agua con gravedad neutra a un plan corporal con mecanismos que permiten al animal respirar aire sin deshidratarse y moverse en tierra es uno de los cambios evolutivos más profundos que se conocen. Los tetrápodos se pueden dividir en cuatro clases: anfibios, reptiles, aves y mamíferos.
| ← tetrapods |
| ||||||||||||||||||
Los tetrápodos marinos son tetrápodos que regresaron de la tierra al mar nuevamente. Los primeros retornos al océano pueden haber ocurrido ya en el período Carbonífero, mientras que otros retornos ocurrieron en fechas tan recientes como el Cenozoico, como en los cetáceos, pinnípedos y varios anfibios modernos. Los anfibios (del griego ambos tipos de vida) viven parte de su vida en el agua y parte en la tierra. En su mayoría requieren agua dulce para reproducirse. Unos pocos habitan en aguas salobres, pero no existen verdaderos anfibios marinos. Sin embargo, ha habido informes de anfibios que han invadido aguas marinas, como una invasión del Mar Negro por parte del híbrido natural Pelophylax esculentus reportada en 2010.
Reptiles
Los reptiles (del latín tardío para arrastrarse o gatear) no tienen una etapa larvaria acuática y, en este sentido, son diferentes a los anfibios. La mayoría de los reptiles son ovíparos, aunque varias especies de escamados son vivíparos, al igual que algunos clados acuáticos extintos: el feto se desarrolla dentro de la madre, contenido en una placenta en lugar de una cáscara de huevo. Como amniotas, los huevos de los reptiles están rodeados de membranas para su protección y transporte, que los adaptan a la reproducción en tierra firme. Muchas de las especies vivíparas alimentan a sus fetos a través de diversas formas de placenta análogas a las de los mamíferos, y algunas brindan cuidado inicial a sus crías.
Algunos reptiles están más estrechamente relacionados con las aves que otros reptiles, y muchos científicos prefieren hacer de Reptilia un grupo monofilético que incluye a las aves. Los reptiles no aviares existentes que habitan o frecuentan el mar incluyen tortugas marinas, serpientes marinas, galápagos, iguanas marinas y cocodrilos de agua salada. Actualmente, de las aproximadamente 12.000 especies y subespecies de reptiles existentes, sólo unas 100 están clasificadas como reptiles marinos.
A excepción de algunas serpientes marinas, la mayoría de los reptiles marinos existentes son ovíparos y necesitan regresar a la tierra para poner sus huevos. Aparte de las tortugas marinas, las especies suelen pasar la mayor parte de su vida en la tierra o cerca de ella, en lugar de en el océano. Las serpientes marinas generalmente prefieren aguas poco profundas cercanas a la tierra, alrededor de islas, especialmente aguas algo protegidas, así como cerca de estuarios. A diferencia de las serpientes terrestres, las serpientes marinas han desarrollado colas aplanadas que les ayudan a nadar.
 Marine iguana
Marine iguana Tortuga marina de cuero
Tortuga marina de cuero Cocodrilo de agua salada
Cocodrilo de agua salada Las serpientes marinas han aplanado colas.
Las serpientes marinas han aplanado colas. El antiguo Ichthyosaurus communis volteretas evolucionadas de forma independiente similares a los delfines.
El antiguo Ichthyosaurus communis volteretas evolucionadas de forma independiente similares a los delfines.

.jpg)
.jpg)


Algunos reptiles marinos extintos, como los ictiosaurios, evolucionaron hasta ser vivíparos y no tenían necesidad de regresar a la tierra. Los ictiosaurios se parecían a los delfines. Aparecieron por primera vez hace unos 245 millones de años y desaparecieron hace unos 90 millones de años. El ancestro terrestre del ictiosaurio no tenía características en su espalda o cola que pudieran haber ayudado en el proceso evolutivo. Sin embargo, el ictiosaurio desarrolló una aleta dorsal y una caudal que mejoraron su capacidad para nadar. El biólogo Stephen Jay Gould dijo que el ictiosaurio era su ejemplo favorito de evolución convergente. Los primeros reptiles marinos surgieron en el Pérmico. Durante el Mesozoico, muchos grupos de reptiles se adaptaron a la vida en los mares, incluidos los ictiosaurios, plesiosaurios, mosasaurios, notosaurios, placodontos, tortugas marinas, talatosaurios y talatosuquios. Los reptiles marinos eran menos numerosos tras la extinción masiva al final del Cretácico.
Pájaros

Las aves marinas están adaptadas a la vida en el entorno marino. A menudo se les llama aves marinas. Si bien las aves marinas varían mucho en estilo de vida, comportamiento y fisiología, a menudo exhiben una sorprendente evolución convergente, ya que los mismos problemas ambientales y nichos de alimentación han dado lugar a adaptaciones similares. Los ejemplos incluyen albatros, pingüinos, alcatraces y alcas.
En general, las aves marinas viven más, se reproducen más tarde y tienen menos crías que las terrestres, pero invierten mucho tiempo en sus crías. La mayoría de las especies anidan en colonias, cuyo tamaño puede variar desde unas pocas docenas de aves hasta millones. Muchas especies son famosas por emprender largas migraciones anuales, cruzar el ecuador o circunnavegar la Tierra en algunos casos. Se alimentan tanto en la superficie del océano como debajo de él, e incluso se alimentan entre sí. Las aves marinas pueden ser altamente pelágicas, costeras o, en algunos casos, pasar una parte del año completamente alejadas del mar. Algunas aves marinas caen en picado desde las alturas, sumergiéndose en el agua dejando estelas parecidas a vapor, similares a las de los aviones de combate. Los alcatraces se sumergen en el agua a hasta 100 kilómetros por hora (60 mph). Tienen sacos de aire debajo de la piel en la cara y el pecho que actúan como plástico de burbujas, amortiguando el impacto con el agua.
 Las gaviotas de arenque europeas atacan escuelas de pastoreo desde arriba.
Las gaviotas de arenque europeas atacan escuelas de pastoreo desde arriba. Pingüino Gentoo nadando bajo el agua
Pingüino Gentoo nadando bajo el agua Los albatros abarcan enormes zonas del océano y algunos círculos del globo.
Los albatros abarcan enormes zonas del océano y algunos círculos del globo. Gannets "divebomb" a alta velocidad
Gannets "divebomb" a alta velocidad




Las primeras aves marinas evolucionaron en el período Cretácico y las familias de aves marinas modernas surgieron en el Paleógeno.
Mamíferos

Los mamíferos (del latín pecho) se caracterizan por la presencia de glándulas mamarias que en las hembras producen leche para alimentar (amamantar) a sus crías. Hay alrededor de 130 especies de mamíferos marinos vivas y recientemente extintas, como focas, delfines, ballenas, manatíes, nutrias marinas y osos polares. No representan un taxón distinto ni una agrupación sistemática, sino que están unificados por su dependencia del medio marino para alimentarse. Tanto los cetáceos como los sirenios son totalmente acuáticos y, por tanto, habitantes obligados del agua. Las focas y los leones marinos son semiacuáticos; Pasan la mayor parte de su tiempo en el agua, pero necesitan regresar a la tierra para realizar actividades importantes como el apareamiento, la reproducción y la muda. Por el contrario, tanto las nutrias como el oso polar están mucho menos adaptados a la vida acuática. Su dieta también varía considerablemente: algunos pueden comer zooplancton; otros pueden comer pescado, calamares, mariscos y algas marinas; y algunos pueden comer otros mamíferos.
En un proceso de evolución convergente, los mamíferos marinos, especialmente los cetáceos como los delfines y las ballenas, volvieron a desarrollar su plan corporal para ser paralelo al plan corporal fusiforme y aerodinámico de los peces pelágicos. Las patas delanteras se convirtieron en aletas y las traseras desaparecieron, reapareció una aleta dorsal y la cola se transformó en una poderosa aleta horizontal. Este plan corporal es una adaptación para ser un depredador activo en un entorno de alta resistencia. Se produjo una convergencia paralela con el ictiosaurio, reptil marino ahora extinto.
![Endangered blue whale, the largest living animal[368]](https://upload.wikimedia.org/wikipedia/commons/thumb/c/cd/Bluewhale2_noaa.jpg/270px-Bluewhale2_noaa.jpg) ballena azul en peligro, el animal vivo más grande
ballena azul en peligro, el animal vivo más grande![The bottlenose dolphin has the highest encephalization of any animal after humans[369]](https://upload.wikimedia.org/wikipedia/commons/thumb/1/10/Tursiops_truncatus_01.jpg/273px-Tursiops_truncatus_01.jpg) El delfín de la nariz de botella tiene la encefalización más alta de cualquier animal después de los humanos
El delfín de la nariz de botella tiene la encefalización más alta de cualquier animal después de los humanos Ballena de Beluga
Ballena de Beluga Dugong pastando sobre margras
Dugong pastando sobre margras Walrus
Walrus Oso polar
Oso polar
![Endangered blue whale, the largest living animal[368]](https://en.wikipedia.org/wiki/File:Bluewhale2_noaa.jpg)
![The bottlenose dolphin has the highest encephalization of any animal after humans[369]](https://en.wikipedia.org/wiki/File:Tursiops_truncatus_01.jpg)



.jpg)
Productoras primarias
(feminine)
Los productores primarios son los organismos autótrofos que elaboran su propio alimento en lugar de comerse otros organismos. Esto significa que los productores primarios se convierten en el punto de partida en la cadena alimentaria para los organismos heterótrofos que se alimentan de otros organismos. Algunos productores primarios marinos son bacterias y arqueas especializadas que son quimiotrofas, que elaboran su propio alimento al reunirse alrededor de respiraderos hidrotermales y filtraciones frías y mediante quimiosíntesis. Sin embargo, la mayor parte de la producción primaria marina proviene de organismos que utilizan la fotosíntesis del dióxido de carbono disuelto en el agua. Este proceso utiliza la energía de la luz solar para convertir agua y dióxido de carbono en azúcares que pueden utilizarse tanto como fuente de energía química como de moléculas orgánicas que se utilizan en los componentes estructurales de las células. Los productores primarios marinos son importantes porque sustentan casi toda la vida animal marina al generar la mayor parte del oxígeno y los alimentos que proporcionan a otros organismos la energía química que necesitan para existir.
Los principales productores primarios marinos son las cianobacterias, las algas y las plantas marinas. Casi todos los seres vivos necesitan el oxígeno liberado como subproducto de la fotosíntesis para llevar a cabo la respiración celular. Además, los productores primarios influyen en los ciclos globales del carbono y el agua. Estabilizan las zonas costeras y pueden proporcionar hábitats para los animales marinos. El término división se ha utilizado tradicionalmente en lugar de filo cuando se habla de productores primarios, pero el Código Internacional de Nomenclatura para algas, hongos y plantas ahora acepta ambos términos como equivalentes.
Cianobacterias

.jpg)
Las cianobacterias fueron los primeros organismos en desarrollar la capacidad de convertir la luz solar en energía química. Forman un filo (división) de bacterias que van desde unicelulares hasta filamentosas e incluyen especies coloniales. Se encuentran en casi todas partes del planeta: en suelos húmedos, en ambientes marinos y de agua dulce, e incluso en rocas antárticas. En particular, algunas especies se presentan como células a la deriva que flotan en el océano y, como tales, estuvieron entre las primeras del fitoplancton.
Los primeros productores primarios que utilizaron la fotosíntesis fueron las cianobacterias oceánicas hace unos 2.300 millones de años. La liberación de oxígeno molecular por las cianobacterias como subproducto de la fotosíntesis indujo cambios globales en el medio ambiente de la Tierra. Debido a que el oxígeno era tóxico para la mayor parte de la vida en la Tierra en ese momento, esto llevó a la casi extinción de los organismos intolerantes al oxígeno, un cambio dramático que reorientó la evolución de las principales especies animales y vegetales.
La diminuta cianobacteria marina Proclorococo, descubierta en 1986, forma hoy parte de la base de la cadena alimentaria oceánica y representa gran parte de la fotosíntesis del océano abierto y aproximadamente el 20% del oxígeno. en la atmósfera terrestre. Es posiblemente el género más abundante de la Tierra: un solo mililitro de agua de mar superficial puede contener 100.000 células o más.
Originalmente, los biólogos clasificaban las cianobacterias como algas y se referían a ellas como "algas verdiazules". La opinión más reciente es que las cianobacterias son bacterias y, por tanto, ni siquiera pertenecen al mismo reino que las algas. Hoy en día, la mayoría de las autoridades excluyen a todos los procariotas y, por tanto, a las cianobacterias de la definición de algas.
Algas
.jpg)
.jpg)


Algas es un término informal para un grupo amplio y diverso de protistas fotosintéticos que no están necesariamente estrechamente relacionados y, por lo tanto, son polifiléticos. Las algas marinas se pueden dividir en seis grupos:
- algas verdes, un grupo informal que contiene alrededor de 8.000 especies reconocidas. Muchas especies viven la mayor parte de sus vidas como células individuales o son filamentosas, mientras que otras forman colonias compuestas de cadenas largas de células, o son algas macroscópicas altamente diferenciadas.
- algas rojas, un phylum (disputado) que contiene alrededor de 7.000 especies reconocidas, principalmente multicelular e incluyendo muchas algas notables.
- algas marrón, una clase que contiene cerca de 2.000 especies reconocidas, principalmente multicelular e incluyendo muchas algas marinas, incluyendo kelp
- diatoms, a (disputed) phylum containing about 100,000 recognized species of mainly unicellular algae. Los átomos generan alrededor del 20 por ciento del oxígeno producido en el planeta cada año, toman más de 6.700 millones de toneladas métricas de silicio cada año de las aguas en las que viven, y aportan casi la mitad del material orgánico que se encuentra en los océanos. Los proyectiles (frustules) de los diatomeas muertos pueden alcanzar hasta media milla de profundidad en el suelo oceánico.
- dinoflagellatos, un phylum de flagelos unicellulares con cerca de 2.000 especies marinas. Se sabe que muchos dinoflagelados son fotosintéticos, pero una gran parte de estos son de hecho mixotropóticos, combinando fotosíntesis con la ingestión de presa (fagotrofia). Algunas especies son endosymbionts de animales marinos y juegan una parte importante en la biología de los arrecifes de coral. Otros preceden a otros protozoos, y algunas formas son parasitarias.
- euglenophytes, un phylum of unicellular flagellates with only a few marine members
A diferencia de las plantas superiores, las algas carecen de raíces, tallos u hojas. Se pueden clasificar por tamaño en microalgas o macroalgas.
LasMicroalgas son los tipos microscópicos de algas, no visibles a simple vista. En su mayoría son especies unicelulares que existen como individuos o en cadenas o grupos, aunque algunas son multicelulares. Las microalgas son componentes importantes de los protistas marinos (que se analizan anteriormente), así como del fitoplancton (que se analiza a continuación). Son muy diversos. Se ha estimado que hay entre 200.000 y 800.000 especies de las cuales se han descrito unas 50.000 especies. Dependiendo de la especie, sus tamaños varían desde unos pocos micrómetros (μm) hasta unos cientos de micrómetros. Están especialmente adaptados a un entorno dominado por fuerzas viscosas.
 Chlamydomonas globosa, una alga verde unicellular con dos flagella sólo visible en la parte inferior izquierda
Chlamydomonas globosa, una alga verde unicellular con dos flagella sólo visible en la parte inferior izquierda![Chlorella vulgaris, a common green microalgae, in endosymbiosis with a ciliate[390]](https://upload.wikimedia.org/wikipedia/commons/thumb/b/b6/%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg/293px-%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg) Clorella vulgaris, una microalga verde común, en endosymbiosis con un ciliate
Clorella vulgaris, una microalga verde común, en endosymbiosis con un ciliate Diatom centrico
Diatom centrico Dinoflagellates
Dinoflagellates
.jpg)
![Chlorella vulgaris, a common green microalgae, in endosymbiosis with a ciliate[390]](https://en.wikipedia.org/wiki/File:%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg)


Macroalgas son los tipos de algas más grandes, multicelulares y más visibles, comúnmente llamadas algas marinas. Las algas marinas suelen crecer en aguas costeras poco profundas, donde están ancladas al fondo marino mediante un anclaje. Las algas que quedan a la deriva pueden llegar a las playas. El quelpo es un gran alga parda que forma grandes bosques submarinos que cubren aproximadamente el 25% de las costas del mundo. Se encuentran entre los ecosistemas más productivos y dinámicos de la Tierra. Algunas algas Sargassum son planctónicas (flotan libremente). Al igual que las microalgas, las macroalgas (algas marinas) son técnicamente protistas marinos ya que no son verdaderas plantas.
 Una algas marinas es una forma macroscópica de algas rojas o marrón o verde.
Una algas marinas es una forma macroscópica de algas rojas o marrón o verde. Sargassum algas es una alga marrón planctónica con vejigas de aire que lo ayudan a flotar.
Sargassum algas es una alga marrón planctónica con vejigas de aire que lo ayudan a flotar. Los peces sargassum son camuflados para vivir entre la deriva Sargassum algas marinas.
Los peces sargassum son camuflados para vivir entre la deriva Sargassum algas marinas.




- macroalgas uniculares (ver también protistas macroscópicos ←)
![The unicellular bubble algae lives in tidal zones. It can have a 4 cm diameter.[393]](https://upload.wikimedia.org/wikipedia/commons/thumb/3/3d/Ventricaria_ventricosa.JPG/248px-Ventricaria_ventricosa.JPG) La alga de burbuja unicellular vive en zonas de marea. Puede tener un diámetro de 4 cm.
La alga de burbuja unicellular vive en zonas de marea. Puede tener un diámetro de 4 cm. La copa de vino de la sirena unicellular son algas en forma de hongos que crecen hasta 10 cm de altura.
La copa de vino de la sirena unicellular son algas en forma de hongos que crecen hasta 10 cm de altura.![Killer algae are single-celled organisms, but look like ferns and grow stalks up to 80 cm long.[394]](https://upload.wikimedia.org/wikipedia/commons/thumb/e/e7/CaulerpaTaxifolia.jpg/316px-CaulerpaTaxifolia.jpg) Las algas asesinas son organismos monocelulares, pero parecen helechos y crecen tallos de hasta 80 cm de largo.
Las algas asesinas son organismos monocelulares, pero parecen helechos y crecen tallos de hasta 80 cm de largo.
![The unicellular bubble algae lives in tidal zones. It can have a 4 cm diameter.[393]](https://en.wikipedia.org/wiki/File:Ventricaria_ventricosa.JPG)

![Killer algae are single-celled organisms, but look like ferns and grow stalks up to 80 cm long.[394]](https://en.wikipedia.org/wiki/File:CaulerpaTaxifolia.jpg)
Los organismos unicelulares suelen ser microscópicos, de menos de una décima de milímetro de largo. Hay excepciones. La copa de sirena, un género de algas verdes subtropicales, es unicelular pero notablemente grande y de forma compleja, con un único núcleo grande, lo que la convierte en un organismo modelo para estudiar la biología celular. Otra alga unicelular, Caulerpa taxifolia, tiene la apariencia de una planta vascular que incluye "hojas" ordenados cuidadosamente en tallos como un helecho. La cría selectiva en acuarios para producir cepas más resistentes dio lugar a una liberación accidental en el Mediterráneo, donde se ha convertido en una especie invasora conocida coloquialmente como alga asesina.
Origen de las plantas

En el Silúrico, parte del fitoplancton evolucionó hasta convertirse en algas rojas, marrones y verdes. Luego, estas algas invadieron la tierra y comenzaron a evolucionar hasta convertirse en las plantas terrestres que conocemos hoy. Posteriormente, en el Cretácico, algunas de estas plantas terrestres regresaron al mar como plantas marinas, como los manglares y las praderas marinas.
Las plantas marinas se pueden encontrar en zonas intermareales y aguas poco profundas, como praderas marinas como la hierba marina y la hierba tortuga, Thalassia. Estas plantas se han adaptado a la alta salinidad del entorno oceánico. La vida vegetal también puede florecer en las aguas salobres de los estuarios, donde pueden crecer manglares, pastos o pastos de playa.
 Mangroves
Mangroves Seagrass meadow
Seagrass meadow![Sea dragons camouflaged to look like floating seaweed live in kelp forests and seagrass meadows.[398]](https://upload.wikimedia.org/wikipedia/commons/thumb/2/2a/Leafy_Sea_Dragon_SA.jpg/120px-Leafy_Sea_Dragon_SA.jpg) Dragones de mar camuflados para parecer como algas flotantes viven en bosques de algas y prados de algas marinas.
Dragones de mar camuflados para parecer como algas flotantes viven en bosques de algas y prados de algas marinas.


![Sea dragons camouflaged to look like floating seaweed live in kelp forests and seagrass meadows.[398]](https://en.wikipedia.org/wiki/File:Leafy_Sea_Dragon_SA.jpg)
La superficie total mundial de bosques de manglares se estimó en 2010 en 134.257 kilómetros cuadrados (51.837 millas cuadradas) (según datos satelitales). La superficie total mundial de praderas de pastos marinos es más difícil de determinar, pero en 2003 se estimó de forma conservadora en 177.000 kilómetros cuadrados (68.000 millas cuadradas).
Los manglares y las praderas marinas proporcionan importantes hábitats de cría para la vida marina, actuando como lugares de escondite y forrajeo para formas larvarias y juveniles de peces e invertebrados más grandes.
Plancton e interacciones tróficas

El plancton (del griego, vagabundos) es un grupo diverso de organismos que viven en la columna de agua de grandes masas de agua pero no pueden nadar contra la corriente. Como resultado, vagan o se dejan llevar por las corrientes. El plancton se define por su nicho ecológico, no por ninguna clasificación filogenética o taxonómica. Son una fuente crucial de alimento para muchos animales marinos, desde peces forrajeros hasta ballenas. El plancton se puede dividir en un componente vegetal y un componente animal.
Fitoplancton
El fitoplancton es el componente vegetal de la comunidad planctónica ("fito" proviene del griego y significa planta). Son autótrofos (se autoalimentan), lo que significa que generan su propio alimento y no necesitan consumir otros organismos.
El fitoplancton está formado principalmente por eucariotas fotosintéticos microscópicos que habitan la capa superior iluminada por el sol en todos los océanos. Necesitan luz solar para poder realizar la fotosíntesis. La mayor parte del fitoplancton son algas unicelulares, pero otros fitoplancton son bacterias y algunos son protistas. Los grupos de fitoplancton incluyen cianobacterias (arriba), diatomeas, varios otros tipos de algas (rojas, verdes, marrones y amarillo verdosas), dinoflagelados, euglenoides, cocolitofóridos, criptomonas, crisofitas, clorofitas, prasinofitas y silicoflagelados. Forman la base de la producción primaria que impulsa la red alimentaria oceánica y representan la mitad de la producción primaria mundial actual, más que los bosques terrestres.


- Phytoplankton
 Phytoplankton son la base de la cadena alimentaria oceánica.
Phytoplankton son la base de la cadena alimentaria oceánica. Phytoplankton viene en muchas formas y tamaños.
Phytoplankton viene en muchas formas y tamaños. Los átomos son uno de los tipos más comunes de fitoplancton.
Los átomos son uno de los tipos más comunes de fitoplancton. Colonial phytoplankton
Colonial phytoplankton


_Various_diatoms.jpg)

 El cyanobacterium Prochloroccus representa gran parte de la producción primaria del océano.
El cyanobacterium Prochloroccus representa gran parte de la producción primaria del océano. Escoria verde cianobacteria lavada en una roca en California
Escoria verde cianobacteria lavada en una roca en California Gyrodinium, uno de los pocos dinoflagelados desnudos que carecen de armadura
Gyrodinium, uno de los pocos dinoflagelados desnudos que carecen de armadura Zooclorolao (verde) viviendo dentro del ciliate Stichotricha secunda
Zooclorolao (verde) viviendo dentro del ciliate Stichotricha secunda



.jpg)

 Coccolithophores nombrado después de la serie documental de la BBC. El Planeta Azul
Coccolithophores nombrado después de la serie documental de la BBC. El Planeta Azul El cocolithophore Emiliania huxleyi
El cocolithophore Emiliania huxleyi Floración de Algae Emiliania huxleyi frente a la costa sur de Inglaterra
Floración de Algae Emiliania huxleyi frente a la costa sur de Inglaterra![Guinardia delicatula, a diatom responsible for algal blooms in the North Sea and the English Channel[407]](https://upload.wikimedia.org/wikipedia/commons/thumb/4/43/Fjouenne_sbrmvr012w_20070924163039_small.jpg/260px-Fjouenne_sbrmvr012w_20070924163039_small.jpg) Guinardia delicatula, un diatom responsable de las floraciones de algas en el Mar del Norte y el Canal Inglés
Guinardia delicatula, un diatom responsable de las floraciones de algas en el Mar del Norte y el Canal Inglés



![Guinardia delicatula, a diatom responsible for algal blooms in the North Sea and the English Channel[407]](https://en.wikipedia.org/wiki/File:Fjouenne_sbrmvr012w_20070924163039_small.jpg)
Zooplancton


El zooplancton es el componente animal de la comunidad planctónica ("zoo" viene del griego y significa animal). Son heterótrofos (se alimentan de otros), lo que significa que no pueden producir sus propios alimentos y, en su lugar, deben consumir otras plantas o animales como alimento. En particular, esto significa que comen fitoplancton.



.jpg)
El zooplancton es generalmente más grande que el fitoplancton, en su mayoría todavía microscópico, pero algunos se pueden ver a simple vista. Muchos protozoos (protistas unicelulares que se alimentan de otra vida microscópica) son zooplancton, incluidos los zooflagelados, los foraminíferos, los radiolarios y algunos dinoflagelados. Otros dinoflagelados son mixotróficos y también podrían clasificarse como fitoplancton; La distinción entre plantas y animales a menudo se rompe en organismos muy pequeños. Otros zooplancton incluyen cnidarios pelágicos, ctenóforos, moluscos, artrópodos y tunicados, así como gusanos flecha planctónicos y gusanos de cerdas.
Los radiolarios son protistas unicelulares con elaboradas conchas de sílice
Microzooplancton: principales herbívoros del plancton
 Los radiólogos vienen en muchas formas.
Los radiólogos vienen en muchas formas. Group of planktic foraminiferans
Group of planktic foraminiferans Los copos comen fitoplancton. Este lleva huevos.
Los copos comen fitoplancton. Este lleva huevos. El dinoflagelado, Protoperidinio extruye un gran velo de alimentación para capturar presa.
El dinoflagelado, Protoperidinio extruye un gran velo de alimentación para capturar presa.




El zooplancton más grande puede ser depredador del zooplancton más pequeño.
Macrozooplancton
 Medusa lunar
Medusa lunar Venus girdle, un citoforo
Venus girdle, un citoforo Arrow gusano
Arrow gusano![Tomopteris, a planktonic segmented worm with unusual yellow bioluminescence[409]](https://upload.wikimedia.org/wikipedia/commons/thumb/4/40/Tomopteriskils.jpg/318px-Tomopteriskils.jpg) Tomopteris, un gusano segmentado planctónico con bioluminiscencia amarilla inusual
Tomopteris, un gusano segmentado planctónico con bioluminiscencia amarilla inusual Anfipodo marino
Anfipodo marino Krill
Krill Pelagic sea cucumber
Pelagic sea cucumber
_Luc_Viatour.jpg)


![Tomopteris, a planktonic segmented worm with unusual yellow bioluminescence[409]](https://en.wikipedia.org/wiki/File:Tomopteriskils.jpg)

.jpg)

Muchos animales marinos comienzan su vida como zooplancton en forma de huevos o larvas, antes de convertirse en adultos. Estos son meroplancticos, es decir, son planctónicos sólo durante una parte de su vida.
- Larvae and juveniles
 Larva salmón recogiendo de su huevo
Larva salmón recogiendo de su huevo Ocean sunfish larva
Ocean sunfish larva Escudo juvenil planctónico
Escudo juvenil planctónico Larva etapa de una langosta espinal
Larva etapa de una langosta espinal




Plancton mixotrófico

.jpg)
Los dinoflagelados suelen ser mixotróficos o vivir en simbiosis con otros organismos.
- Mixoplankton
 Tintinnid ciliate Favella
Tintinnid ciliate Favella Euglena mutabilis, un bandellate fotosintético
Euglena mutabilis, un bandellate fotosintético Noctiluca scintillans, un dinoflagelato de bioluminiscencia
Noctiluca scintillans, un dinoflagelato de bioluminiscencia


Algunos dinoflagelados son bioluminiscentes. Por la noche, el agua del océano puede iluminarse internamente y brillar con una luz azul debido a estos dinoflagelados. Los dinoflagelados bioluminiscentes poseen centelleos, cuerpos citoplasmáticos individuales que contienen dinoflagelados luciferasa, la principal enzima implicada en la luminiscencia. La luminiscencia, a veces llamada la fosforescencia del mar, se produce como breves destellos azules (0,1 segundos) o chispas cuando se estimulan centelleos individuales, generalmente por perturbaciones mecánicas de, por ejemplo, un barco o un nadador o navegar.
Red alimentaria marina

En comparación con los ambientes terrestres, los ambientes marinos tienen pirámides de biomasa que están invertidas en la base. En particular, la biomasa de los consumidores (copépodos, krill, camarones, peces forrajeros) es mayor que la biomasa de los productores primarios. Esto sucede porque los productores primarios del océano son pequeños fitoplancton que tienden a ser estrategas r que crecen y se reproducen rápidamente, por lo que una masa pequeña puede tener una tasa rápida de producción primaria. Por el contrario, los productores primarios terrestres, como los bosques maduros, suelen ser estrategas K que crecen y se reproducen lentamente, por lo que se necesita una masa mucho mayor para lograr la misma tasa de producción primaria.
Debido a esta inversión, es el zooplancton el que constituye la mayor parte de la biomasa animal marina. Como consumidores primarios, son el vínculo crucial entre los productores primarios (principalmente fitoplancton) y el resto de la red alimentaria marina (consumidores secundarios).
Si el fitoplancton muere antes de ser comido, desciende a través de la zona eufótica como parte de la nieve marina y se asienta en las profundidades del mar. De esta manera, el fitoplancton secuestra alrededor de 2 mil millones de toneladas de dióxido de carbono en el océano cada año, lo que hace que el océano se convierta en un sumidero de dióxido de carbono que contiene alrededor del 90% de todo el carbono secuestrado.
En 2010, los investigadores descubrieron que las ballenas transportan nutrientes desde las profundidades del océano a la superficie mediante un proceso que llamaron bomba de ballena. Las ballenas se alimentan en niveles más profundos del océano donde se encuentra el krill, pero regresan regularmente a la superficie para respirar. Allí las ballenas defecan un líquido rico en nitrógeno y hierro. En lugar de hundirse, el líquido permanece en la superficie donde el fitoplancton lo consume. En el golfo de Maine, la bomba de ballenas proporciona más nitrógeno que los ríos.
Otras interacciones
Ciclos biogeoquímicos

En su conjunto, los océanos forman un único sistema marino donde el agua, el "disolvente universal" – disuelve nutrientes y sustancias que contienen elementos como oxígeno, carbono, nitrógeno y fósforo. Estas sustancias se ciclan y reciclan sin cesar, se combinan químicamente y luego se descomponen nuevamente, se disuelven y luego se precipitan o evaporan, se importan y se exportan de regreso a la tierra, la atmósfera y el fondo del océano. Impulsados tanto por la actividad biológica de los organismos marinos como por las acciones naturales del sol y las mareas y movimientos dentro de la corteza terrestre, estos son los ciclos biogeoquímicos marinos.
![Marine carbon cycle[421]](https://upload.wikimedia.org/wikipedia/commons/thumb/0/03/OceanCarbonCycle.jpg/223px-OceanCarbonCycle.jpg) Ciclo de carbono marino
Ciclo de carbono marino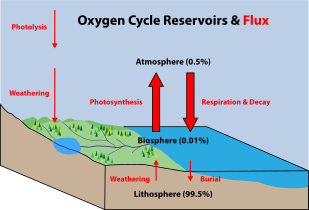 Ciclo de oxígeno
Ciclo de oxígeno
![Marine carbon cycle[421]](https://en.wikipedia.org/wiki/File:OceanCarbonCycle.jpg)

 Ciclo de nitrógeno marino
Ciclo de nitrógeno marino Ciclo de fósforo marino
Ciclo de fósforo marino


Sedimentos y exudados biogénicos
.jpg)
Los sedimentos en el fondo del océano tienen dos orígenes principales, terrígenos y biógenos. Los sedimentos terrígenos representan alrededor del 45% del total de sedimentos marinos y se originan en la erosión de las rocas terrestres, transportadas por ríos y escorrentías terrestres, polvo transportado por el viento, volcanes o trituración de glaciares.
Los sedimentos biógenos representan el otro 55% del sedimento total y se originan en los restos esqueléticos de protistas marinos (organismos unicelulares de plancton y bentos). También pueden estar presentes cantidades mucho más pequeñas de minerales precipitados y polvo meteórico. Modo, en el contexto de un sedimento marino, no se refiere a la consistencia del sedimento sino a su origen biológico. El término exudado fue utilizado originalmente por John Murray, el "padre de la oceanografía moderna", quien propuso el término exudado radiolario para los depósitos de sílice de las conchas de radiolarios que salieron a la superficie durante el Challenger. Expedición. Un lodo biogénico es un sedimento pelágico que contiene al menos un 30 por ciento de restos esqueléticos de organismos marinos.
Principales tipos de ooze biogénico | |||||||
|---|---|---|---|---|---|---|---|
| Tipo | mineral formas | protista responsable | nombre Esqueleto | descripción | |||
| Ooze piadoso | SiO2 quartz cristal opal masticar | diatomeas |  | frustule | Los diatomeas individuales varían en tamaño de 0.002 a 0.2 mm. | ||
| radiólogos | .jpg) | Esqueleto | Los radiólogos son protozoos con diámetros típicamente entre 0,1 y 0,2 mm que producen esqueletos minerales intrincados, generalmente hechos de sílice | ||||
| Ooze calcáreo | CaCO3 calcite aragonita piedra caliza tiza | foraminiferans | | prueba | Hay unas 10.000 especies vivas de foraminiferanos, generalmente menores de 1 mm de tamaño. | ||
| cocolithophores |  | cocolith | Los cocolithophores son células esféricas generalmente menos de 0,1 mm de ancho, encerradas por placas calcáreas llamadas cocolitos. Los cocolitos son microfisiles importantes. Son la mayor fuente mundial de carbonato de calcio bigénico y hacen contribuciones significativas al ciclo mundial del carbono. Son el principal componente de depósitos de tiza como los acantilados blancos de Dover. | ||||
 Un esqueleto mineral elaborado de un radiolarian hecho de sílice.
Un esqueleto mineral elaborado de un radiolarian hecho de sílice. Diatoms, componentes principales del plancton marino, también tienen esqueletos de silica llamados frustules.
Diatoms, componentes principales del plancton marino, también tienen esqueletos de silica llamados frustules. Cocolitofores tienen placas o escamas hechas con carbonato de calcio llamado cocolitos
Cocolitofores tienen placas o escamas hechas con carbonato de calcio llamado cocolitos Prueba calcificada de un foraminiferan plancético
Prueba calcificada de un foraminiferan plancético
,_Haeckel_(28187768550).jpg)



 A diatom microfossil from 40 million years ago
A diatom microfossil from 40 million years ago La tierra diatomácea es una roca sedimentaria suave, silícea, formada por microfosils en forma de frustules (piezas) de diatomeas de células individuales (clic para magnificar).
La tierra diatomácea es una roca sedimentaria suave, silícea, formada por microfosils en forma de frustules (piezas) de diatomeas de células individuales (clic para magnificar). Ilustración de una Globigerina Ooze
Ilustración de una Globigerina Ooze Shells (tests), usualmente hecho de carbonato de calcio, de un ooze foraminiferal en el fondo oceánico
Shells (tests), usualmente hecho de carbonato de calcio, de un ooze foraminiferal en el fondo oceánico




Interacciones terrestres

Las interacciones terrestres impactan la vida marina de muchas maneras. Las costas suelen tener plataformas continentales que se extienden a cierta distancia de la costa. Estos proporcionan extensas aguas poco profundas iluminadas por el sol hasta el fondo marino, lo que permite la fotosíntesis y propicia hábitats para praderas de pastos marinos, arrecifes de coral, bosques de algas marinas y otras formas de vida bentónica. Más lejos de la costa, la plataforma continental se inclina hacia aguas profundas. El viento que sopla en la superficie del océano o las corrientes oceánicas profundas pueden provocar que aguas frías y ricas en nutrientes provenientes de profundidades abisales asciendan por los taludes continentales. Esto puede dar lugar a surgencias a lo largo de los bordes exteriores de las plataformas continentales, proporcionando las condiciones para la proliferación de fitoplancton.
El agua evaporada por el sol desde la superficie del océano puede precipitarse en la tierra y eventualmente regresar al océano como escorrentía o descarga de los ríos, enriquecida con nutrientes y contaminantes. A medida que los ríos desembocan en los estuarios, el agua dulce se mezcla con el agua salada y se vuelve salobre. Esto proporciona otro hábitat de aguas poco profundas donde prosperan los bosques de manglares y los peces de estuario. En general, la vida en los lagos interiores puede evolucionar con mayor diversidad que la que ocurre en el mar, porque los hábitats de agua dulce son en sí mismos diversos y compartimentados de una manera que los hábitats marinos no lo son. Algunas formas de vida acuática, como el salmón y las anguilas, migran de un lado a otro entre hábitats marinos y de agua dulce. Estas migraciones pueden resultar en intercambios de patógenos y tener impactos en la forma en que evoluciona la vida en el océano.
Impactos antropogénicos

Las actividades humanas afectan la vida marina y los hábitats marinos a través de la sobrepesca, la contaminación, la acidificación y la introducción de especies invasoras. Estos impactan los ecosistemas marinos y las redes alimentarias y pueden tener consecuencias aún no reconocidas para la biodiversidad y la continuación de las formas de vida marina.
Eventos de biodiversidad y extinción


La biodiversidad es el resultado de más de tres mil millones de años de evolución. Hasta hace aproximadamente 600 millones de años, toda la vida estaba formada por arqueas, bacterias, protozoos y organismos unicelulares similares. La historia de la biodiversidad durante el Fanerozoico (los últimos 540 millones de años) comienza con un rápido crecimiento durante la explosión del Cámbrico, un período durante el cual aparecieron por primera vez casi todos los filos de organismos multicelulares. Durante los siguientes 400 millones de años aproximadamente, la diversidad de invertebrados mostró poca tendencia general y la diversidad de vertebrados muestra una tendencia exponencial general.
Sin embargo, se estima que más del 99 por ciento de todas las especies que alguna vez vivieron en la Tierra, es decir, más de cinco mil millones de especies, están extintas. Estas extinciones ocurren a un ritmo desigual. El espectacular aumento de la diversidad ha estado marcado por pérdidas periódicas y masivas de diversidad clasificadas como eventos de extinción masiva. Los eventos de extinción masiva ocurren cuando la vida sufre un declive global precipitado. La mayor diversidad y biomasa de la Tierra se encuentra entre los microorganismos, que son difíciles de medir. Por lo tanto, los eventos de extinción registrados se basan en los cambios más fácilmente observables en la diversidad y abundancia de organismos multicelulares más grandes, en lugar de en la diversidad y abundancia total de la vida. Los fósiles marinos se utilizan principalmente para medir las tasas de extinción debido a su registro fósil y rango estratigráfico superiores en comparación con los organismos terrestres.
Según el registro fósil, la tasa de extinciones en la Tierra es de aproximadamente dos a cinco familias taxonómicas de animales marinos cada millón de años. El Gran Evento de Oxigenación fue quizás el primer gran evento de extinción. Desde la explosión del Cámbrico, cinco grandes extinciones masivas han excedido significativamente la tasa de extinción previa. El peor fue el evento de extinción del Pérmico-Triásico, hace 251 millones de años. En general, se estima que las cinco grandes extinciones masivas del Fanerozoico (los últimos 540 millones de años) acabaron con más del 40% de los géneros marinos y probablemente más del 70% de las especies marinas. La actual extinción del Holoceno causada por la actividad humana, y ahora denominada "sexta extinción", puede resultar, en última instancia, más devastadora.
Investigando y explorando la vida marina
Investigación y estudio
Para realizar investigaciones y enriquecer el conocimiento sobre la vida marina, los científicos utilizan varios métodos para alcanzar y explorar las profundidades del océano. Para ello se utilizan varios instrumentos y vehículos de alta tecnología.
- Vehículos submarinos autónomos (UA)- Los robots submarinos solían explorar el océano. Los AUV son robots independientes y pueden explorar sin tripulación. Son liberados de un barco y son operados desde la superficie.
- Vehículos de remolque profundo (DTVs) remolcados tras buques de investigación, ofreciendo una alternativa más sencilla a vehículos submarinos más avanzados. Sirven como plataformas versátiles para implementar instrumentos oceanográficos para medir diversos parámetros oceánicos, con modelos específicos como el DTV BRIDGET utilizado para estudiar ciruelas de ventilación hidrotermal moviéndose cerca del suelo oceánico.
- Manned Submersibles- un vehículo submarino tripulado utilizado para explorar, experimentar y es a menudo utilizado por el ejército.
- Naves de investigación (R/Vs)- un barco o barco utilizado para realizar investigaciones durante un período de tiempo. Es capaz de transportar una amplia gama de equipos de muestreo y reconocimiento. Los buques de investigación suelen tener espacio de laboratorio a bordo, lo que permite a los investigadores analizar rápidamente los materiales recogidos durante los cruceros.
- Vehículos operados a distancia (ROV)- vehículos no tripulados. capaz de alcanzar mayores profundidades bajo el agua para recoger una mayor variedad de información.