
Topoisomerasa
Descubrimiento
La primera topoisomerasa de ADN fue descubierta en bacterias por James Wang en 1971 e inicialmente se denominó proteína ω (omega); ahora se llama Escherichia coli (E. coli) topoisomerasa I (topo I) y es un representante de la familia de enzimas tipo IA. Posteriormente, James Champoux y Renato Dulbecco encontraron una actividad similar en células eucariotas (hígado de rata); la enzima responsable, eucariota topo I, tiene un mecanismo distinto y es representativa de la familia tipo IB. La primera topoisomerasa tipo II que se descubrió fue la ADN girasa de bacterias, por Martin Gellert y colaboradores en 1976, y también caracterizada por Nicholas Cozzarelli y colaboradores. La ADN girasa cataliza la introducción de superenrollamientos negativos en el ADN y es la única enzima de tipo II que hace esto, todas las demás catalizan la relajación del ADN. Las enzimas de tipo II se diferencian mecánicamente de las de tipo I en que dependen de ATP y escinden transitoriamente ambas cadenas de ADN en lugar de solo una. Las topoisomerasas de tipo II se identificaron posteriormente a partir de virus bacterianos y eucariotas. Los códigos EC de Topo son los siguientes: independiente de ATP (tipo I), EC 5.6.2.1; Dependiente de ATP (tipo II): EC 5.6.2.2. La excepción entre las topoisomerasas de tipo I, la girasa inversa, que contiene un dominio helicasa (EC 3.6.4.12) e introduce un superenrollamiento positivo de forma dependiente de ATP. Por lo tanto, es la única topoisomerasa tipo I clasificada como EC 5.6.2.2 (Tabla 1).
Topología de ADN
La estructura de doble hélice del ADN implica el entrelazamiento de las dos cadenas de polinucleótidos entre sí, lo que potencialmente da lugar a problemas topológicos. La topología del ADN se refiere al cruce de las dos cadenas de ADN que altera el giro de la doble hélice y da lugar a conformaciones terciarias del ADN, como superenrollamientos, nudos y catenanas. Los posibles problemas topológicos asociados con la estructura de doble hélice del ADN se reconocieron poco después de que James Watson, Francis Crick y Rosalind Franklin aclararan su estructura por primera vez en 1953 y se desarrollaron aún más con el trabajo de Max Delbruck y John Cairns. El ADN bicatenario circular cerrado se puede describir mediante 3 parámetros: número de enlace (Lk), torsión (Tw) y retorcimiento (Wr) (Fig. 1). Donde Lk se refiere al número de veces que se unen las dos hebras, Tw se refiere al número de vueltas helicoidales en el ADN, medido en relación con el eje helicoidal, y Wr cuantifica el enrollamiento de la trayectoria de la hélice del ADN en el espacio y, a menudo, es equiparado con 'superenrollamiento'.
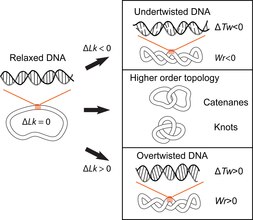
Los 3 parámetros están relacionados de la siguiente manera: Lk = Tw +Wr. Esta es una identidad matemática obtenida originalmente por Călugăreanu en 1959 y se conoce como el teorema de Călugăreanu o Călugăreanu-White-Fuller. Lk no puede alterarse sin romper una o ambas hebras de la hélice; Tw y Wr son interconvertibles y dependen de las condiciones de la solución. Superenrollamiento es un término vernáculo para el ADN con una diferencia de enlace distinta de cero, más correctamente denominada diferencia de enlace específica (σ = ΔLk/Lk0, donde Lk0 es el número de enlace medio del círculo de ADN relajado). Se dice que el ADN está superenrollado positivamente si Lk es mayor que Lk0 para el estado relajado (Lk-Lko = ΔLk, ΔLk>0); eso significa que Tw y/o Wr aumentan con respecto a la molécula relajada. Por el contrario, el ADN se superenrolla negativamente si la Lk de la molécula es menor que la Lk0 (ΔLk<0).
Las consecuencias de las perturbaciones topológicas en el ADN se ejemplifican en la replicación del ADN durante la cual se separan las hebras del dúplex; esta separación conduce a la formación de superenrollamientos positivos (sobreenrollamiento o sobretorsión del ADN) por delante de la horquilla de replicación y el entrelazamiento de las hebras hijas (precatenanos) por detrás (Fig. 2). Si los superenrollamientos positivos no se relajan, se impide la progresión de la horquilla de replicación, mientras que el hecho de no desvincular las hebras hijas impide la segregación del genoma, que es necesaria para la división celular. La transcripción por la ARN polimerasa también genera un superenrollamiento positivo por delante y un superenrollamiento negativo por detrás del complejo transcripcional (Fig. 2). Este efecto se conoce como el modelo de dominio doble superenrollado, como lo describieron Leroy Liu y James Wang en 1987. Estas perturbaciones topológicas deben resolverse para que prosiga el metabolismo del ADN, lo que permite que la célula se replique, transcriba y divida el genoma de manera eficiente para permitir que la célula división y vitalidad. Los nudos en el ADN se pueden encontrar en bacteriófagos y como productos de reacciones de recombinación. En general, los nudos en el ADN son perjudiciales y deben eliminarse (mediante topoisomerasas). Los catenanos de ADN se forman tras la replicación de moléculas circulares y deben resolverse mediante topoisomerasas o recombinasas para permitir la separación adecuada de las moléculas hijas durante la división celular. Además de los aspectos perjudiciales de la topología del ADN que requieren resolución, también existen aspectos beneficiosos. Por ejemplo, la replicación del plásmido requiere un superenrollamiento negativo del origen, lo que facilita la fusión local y expone el ADN monocatenario necesario para iniciar la replicación. De manera similar, el inicio de la replicación desde el origen bacteriano principal oriC también requiere un superenrollamiento negativo. Además, la compactación de la E. coli se logra en parte por superenrollamiento negativo.
Tipos
Las topoisomerasas de ADN son enzimas que han evolucionado para resolver problemas topológicos en el ADN (Tabla 2). Lo hacen a través de la rotura transitoria de una o ambas cadenas de ADN. Esto ha llevado a clasificar los topos en dos tipos: tipo I, que catalizan reacciones que implican roturas transitorias de una sola cadena, y tipo II, que catalizan reacciones que implican roturas transitorias de doble cadena (fig. 3; tabla 2). Existen subtipos dentro de estas clasificaciones.

Tipo I
Estas enzimas catalizan cambios en la topología del ADN a través de rupturas transitorias de una sola hebra en el ADN. Las reacciones pueden ocurrir tanto en sustratos de ADN monocatenarios como bicatenarios y pueden proceder a través de un mecanismo 'giratorio' o 'pasaje de hilo' mecanismo (Fig. 3). La gama de reacciones incluye: relajación del superenrollamiento del ADN, desanudamiento de los círculos monocatenarios y decatenación, siempre que al menos una pareja tenga una región monocatenaria. En el caso de la enzima archaeal, la girasa inversa, es posible un superenrollamiento positivo del ADN.
Tipo IA
Los tipos IA son monoméricos y se unen a segmentos de ADN monocatenarios. Introducen una ruptura monocatenaria transitoria a través de la formación de un enlace tirosil-fosfato entre una tirosina en la enzima y un 5'-fosfato en el ADN. El segmento de ADN dentro del cual se produce la ruptura se denomina 'puerta'. o segmento G, y su escisión permite el paso de otro segmento de ADN, el 'transporte' o segmento en T, para pasar a través de un 'pasaje de hebra' proceso. A esto le sigue la ligadura del segmento G. Para que se produzca el paso de la hebra, la topo IA debe sufrir un cambio conformacional para abrir la puerta del ADN y permitir la transferencia del segmento T. Durante una reacción de relajación del ADN, este proceso cambia el número de enlaces del ADN en +/-1 (Fig. 4). Los ejemplos de topoisomerasas de tipo IA incluyen procariotas topo I y III, eucariotas topo IIIα y IIIβ y la enzima arqueal inversa girasa. La girasa inversa, que ocurre en las arqueas termófilas, comprende un topo tipo IA acoplado a una helicasa, y es la única enzima conocida que puede introducir superenrollamientos positivos en el ADN. El gen que codifica la girasa inversa también se encuentra en algunos grupos de bacterias termófilas, donde probablemente se transfirió mediante transferencia horizontal de genes desde Archaea.

Tipo IB
Las topoisomerasas de tipo IB catalizan reacciones que implican roturas monocatenarias transitorias en el ADN a través de la formación de un enlace tirosil-fosfato entre una tirosina en la enzima y un 3'-fosfato en el ADN. En lugar de utilizar un mecanismo de paso de hebra, estas enzimas operan a través de un mecanismo de 'giro'. o 'rotación controlada' de la hebra escindida alrededor de la hebra intacta. Este mecanismo de rotación controlada se describió por primera vez para Vaccinia topo I y permite la rotación del ADN del extremo libre alrededor de la hebra intacta, siendo controlada la velocidad por la 'fricción'. dentro de la cavidad de la enzima, antes de volver a ligar la muesca (Fig. 3). Esto da como resultado un cambio variable del número de enlaces por evento de escisión y religación. Este mecanismo es distinto del de las enzimas de tipo IA, y los dos grupos de enzimas no están relacionados estructural ni evolutivamente. Los ejemplos de topoisomerasas de tipo IB incluyen topo I nuclear y mitocondrial eucariota además de topo I viral, aunque se han identificado en los tres dominios de la vida.
Tipo CI
Las topoisomerasas de tipo IC comparten un mecanismo similar a las enzimas de tipo IB, pero son estructuralmente distintas. El único representante es topo V, que se encuentra en el hipertermófilo Methanopyrus kandleri.
Tipo II
Las topoisomerasas de tipo II catalizan cambios en la topología del ADN a través de roturas transitorias de doble cadena en el ADN. Las reacciones ocurren en sustratos de ADN de doble cadena y proceden a través de un mecanismo de paso de cadena (Fig. 5). La gama de reacciones incluye la relajación del ADN, el superenrollamiento del ADN, el desanudamiento y la decatenación. Mientras que todas las topoisomerasas de tipo II pueden catalizar la relajación del ADN, la girasa, una topoisomerasa bacteriana arquetípica, también puede introducir superenrollamientos negativos. A diferencia de las topoisomerasas de tipo I que son generalmente monoméricas, las topoisomerasas de tipo II son homodímeros o heterotetrámeros. Se clasifican en dos subtipos según consideraciones evolutivas, estructurales y mecanicistas. El mecanismo general de paso de cadena para el topos tipo II comienza con la unión de un dúplex de ADN, denominado segmento de puerta (segmento G), en la puerta de ADN. Otro dúplex, denominado segmento de transporte (segmento T), es capturado por una abrazadera operada por ATP y pasa a través de una ruptura transitoria en el segmento G, que involucra enlaces de fosfotirosina 5ʹ en ambas hebras, antes de ser liberado a través de la puerta C. y se vuelve a ligar el segmento G (Fig. 5). El recambio enzimático requiere la unión e hidrólisis de ATP.

Tipo IIA
Las topoisomerasas de tipo IIA catalizan rupturas transitorias de doble cadena en el ADN a través de la formación de enlaces tirosil-fosfato entre tirosinas en la enzima (una en cada subunidad) y 5'-fosfatos escalonados por 4 bases en cadenas opuestas de ADN. La reacción de paso de la hebra puede ser intra o intermolecular (Fig. 5), lo que permite cambios en el superenrollamiento y el anudado o la desconexión, respectivamente. Este proceso cambia el número de enlace del ADN en +/-2. Los ejemplos de topoisomerasas de tipo IIA incluyen topo IIα y topo IIβ eucariotas, además de girasa bacteriana y topo IV. La ADN girasa se ajusta al mismo mecanismo de paso de doble cadena que otras enzimas de tipo II, pero tiene características únicas relacionadas con su capacidad para introducir superenrollamientos negativos en el ADN. El segmento G es parte de una pieza de ADN mucho más larga (>100 pb) que se envuelve alrededor de la enzima, un brazo del cual forma el segmento T que pasa a través de la rotura de doble cadena (Fig. 5). En el caso de la girasa, una cantidad sustancial de la energía libre de la hidrólisis de ATP se traduce en estrés torsional en el ADN, es decir, el superenrollamiento es un proceso que requiere energía. Además, en ausencia de ATP, la girasa puede eliminar superenrollamientos negativos en una reacción de relajación del ADN más lenta.
Tipo IIB
El tipo IIB también cataliza rupturas transitorias de doble cadena a través de la formación de enlaces tirosil-fosfato entre tirosinas en la enzima y 5'-fosfatos en cadenas opuestas del ADN, pero en el caso de las enzimas IIB, las rupturas de doble cadena tienen un escalonamiento de 2 bases. Las enzimas de tipo IIB muestran importantes diferencias estructurales, pero están relacionadas evolutivamente con las enzimas de tipo IIA. Estas diferencias incluyen la falta de una de las "puertas" de la proteína. (la puerta C) (Fig. 5). Encontrados originalmente en arqueas, también se han encontrado en eucariotas y, en particular, en plantas; los ejemplos incluyen topo VI y topo VIII. Topo VI es la enzima mejor estudiada de este subtipo y se cree que es una decatenasa preferencial.
Como dianas farmacológicas
Para los no especialistas, quizás el aspecto más importante de las topoisomerasas es su función como dianas farmacológicas para la quimioterapia antibacteriana y anticancerígena; varios medicamentos antibacterianos y anticancerígenos dirigidos contra la topoisomerasa se incluyen en la Lista modelo de medicamentos esenciales de la Organización Mundial de la Salud de 2019. La razón de esta prominencia es que sus reacciones proceden a través de roturas transitorias en el ADN que, si se estabilizan mediante la unión de fármacos, pueden provocar la muerte celular debido a la generación de roturas tóxicas monocatenarias o bicatenarias en el ADN genómico. La mayoría de los fármacos topodirigidos actúan de esta manera, es decir, estabilizan el intermedio de escisión covalente enzima-ADN.
Compuestos antibacterianos
Aunque los topos tipo I, como el topo I bacteriano, son objetivos antibióticos viables, actualmente no hay compuestos en uso clínico que se dirijan a estas enzimas. Sin embargo, las enzimas de tipo II, la ADN girasa y la ADN topoisomerasa IV, han disfrutado de un enorme éxito como dianas para los antibióticos de fluoroquinolona ampliamente utilizados (Fig. 6).
Fluoroquinolonas (FQ)
Los compuestos antibacterianos de quinolona se desarrollaron por primera vez en la década de 1960 y han estado en uso clínico desde la década de 1980. Los derivados de FQ, como ciprofloxacino, levofloxacino y moxifloxacino (fig. 6), han tenido un gran éxito. Estos compuestos funcionan interactuando con su objetivo (girasa o topo IV) y el ADN en el sitio de escisión para estabilizar el intermedio de escisión covalente de ADN-proteína. Específicamente, se intercalan en el ADN y previenen el paso de religación del ADN de la reacción de la topoisomerasa (Fig. 5). Este es un mecanismo de inhibición altamente efectivo que también es utilizado por varios medicamentos contra el cáncer dirigidos contra la topoisomerasa. A pesar de su éxito espectacular, la resistencia a las FQ es un problema grave. Una variedad de otros compuestos, como las quinazolinedionas y las imidazolpirazinonas, funcionan de manera similar y se espera que algunos de ellos reemplacen a las FQ en el futuro.
Aminocumarinas
Las aminocumarinas (fig. 6), como la novobiocina, la clorobiocina y la cumermicina A1, son productos naturales de Streptomyces que inhiben la reacción ATPasa de la girasa y la topo IV. Aunque pueden ser muy potentes contra su objetivo, sufren problemas de permeabilidad y toxicidad y, por lo tanto, no han disfrutado del nivel de éxito clínico de las FQ.
Inhibidores proteináceos
Hay una serie de proteínas inhibidoras de la girasa, incluidas las toxinas bacterianas CcdB, MccB17 y ParE, que estabilizan el complejo de escisión, de manera similar a las FQ. Aunque estas proteínas no son viables como antibacterianos, su modo de acción podría inspirar el desarrollo de nuevos compuestos antibacterianos. Otros inhibidores proteicos de la girasa evitan que la topoisomerasa se una al ADN en lugar de estabilizar los complejos de escisión. Estos incluyen YacG y proteínas repetidas de pentapéptidos, como QnrB1 y MfpA; estos inhibidores de proteínas también confieren resistencia a las fluoroquinolonas.

Compuestos contra el cáncer
Tanto la topo I humana como la topo II (ambas isoformas α y β) pueden ser el objetivo de la quimioterapia contra el cáncer (Fig. 7). La mayoría de estos compuestos actúan de manera similar a las FQ, es decir, estabilizando el complejo de escisión covalente ADN-proteína; por esto se les conoce como venenos de topoisomerasa, distintos de los inhibidores catalíticos. Varios inhibidores de la topoisomerasa humana están incluidos en la Lista de medicamentos esenciales de la Organización Mundial de la Salud.
Camptotecina (CPT)
La camptotecina (Fig. 7), originalmente derivada del árbol Camptotheca acuminata, se dirige a la topo I humana y derivados como el topotecán y el irinotecán se usan ampliamente en la quimioterapia contra el cáncer. La camptotecina y sus derivados actúan estabilizando el complejo de escisión topo I, evitando la religación de la mella mediada por proteína en el ADN. Estos inhibidores interfaciales se estabilizan mediante interacciones de apilamiento con el ADN cortado y los enlaces de hidrógeno con la enzima. Aunque los derivados de CPT estabilizan un complejo de escisión monocatenario, se cree que las colisiones posteriores con la maquinaria de replicación o transcripción generan roturas tóxicas de ADN bicatenario. Estos compuestos se utilizan como terapias de primera o segunda línea para tratar cánceres, incluidos el colorrectal, el de ovario, el de pulmón, el de mama y el de cuello uterino. Sin embargo, los derivados de CPT sufren limitaciones asociadas con la toxicidad y vidas medias terapéuticas limitadas debido a la inestabilidad química. Los nuevos inhibidores de la topo I, las indenoisoquinolinas y las fluoroindenoisoquinolinas, superan las limitaciones de los derivados de la CPT y actualmente se encuentran en ensayos clínicos.
Etopósido (VP-16)
El etopósido (Fig. 7) y su pariente cercano el tenipósido (VM-26) son derivados de la epipodofilotoxina obtenidos del rizoma de la mandrágora silvestre que se dirigen a la topo II al estabilizar el complejo de escisión covalente y prevenir la religación del ADN escindido. Por lo general, se usan junto con otros medicamentos de quimioterapia para tratar cánceres, incluidos los tumores testiculares, el cáncer de pulmón de células pequeñas y la leucemia. El tratamiento con etopósido puede dar lugar a leucemias secundarias derivadas de translocaciones genómicas específicas, que afectan principalmente a topo IIβ.
Doxorrubicina

La doxorrubicina (Fig. 7) y los derivados relacionados daunorrubicina, epirrubicina e idarrubicina son antraciclinas obtenidas de la bacteria Streptomyces que se dirigen a la topo II humana, estabilizando el complejo de escisión de manera similar a otras topoisomerasas venenos La mitoxantrona es una antracenodiona sintética química y funcionalmente similar a las antraciclinas. Las antraciclinas fueron los primeros inhibidores de la topoisomerasa utilizados para tratar el cáncer y se encuentran entre los tratamientos más utilizados y efectivos para una amplia gama de cánceres, incluidos el cáncer de mama, el linfoma, las leucemias, los carcinomas, los sarcomas y otros tumores. Estos compuestos son agentes intercalantes del ADN y, como tales, pueden afectar una amplia gama de procesos del ADN celular además de envenenar específicamente a topo II. La citotoxicidad adicional proviene de reacciones redox que involucran antraciclinas que generan especies reactivas de oxígeno. La generación de oxígeno reactivo, junto con el envenenamiento de topo IIβ, da como resultado la cardiotoxicidad limitante de la dosis de las antraciclinas.
Merbarona
La merbarona es un derivado del ácido tiobarbitúrico y el dexrazoxano (ICRF-187), uno de varios derivados de bisdioxopiperazina relacionados (Fig. 7), son ejemplos de inhibidores catalíticos de topo II, es decir, evitan que se complete el ciclo catalítico de topo II. pero no estabilizan el complejo de escisión del ADN. Si bien estos inhibidores catalíticos exhiben citotoxicidad y se han probado en ensayos clínicos, actualmente no se usan clínicamente para la terapia contra el cáncer. Sin embargo, el dexrazoxano, que bloquea la hidrólisis de ATP por topo II, se usa para prevenir la cardiotoxicidad asociada con las antraciclinas.
| Topoisomerase | Tipo de subfamilia | Función | Multimericity | Metal dependence | ATP dependence | ¿Uno o doble escote? | polaridad salvaje | Cambio en el número de enlace (L) |
|---|---|---|---|---|---|---|---|---|
| Topoisomerasa I
()E. coli) | Tipo IA | Quita (-), pero no (+) supercoils. Evita la supercoiling excesiva del genoma, y apoya la transcripción | Monomer | Sí (Mg2+) | No | SS | ||
| Topoisomerasa III
()E. coli) | Elimina (-), pero no (+) supercoils; función superpuesta con topoisomerasa IV | |||||||
| Topoisomerasa IIIα
()H. sapiens) | Elimina (-), pero no (+) supercoils; ayuda en el desenlace de precatenanías en la replicación del ADN celular; puede catalizar el nudo, desenganchado, e interrelacionado de círculos monoplazados, así como el nudo, desenganchado, catenación, y la decatenación de círculos de ADN dúplex accionados o nudos | |||||||
| Topoisomerasa IIIβ
()H. sapiens) | Se ha demostrado que es una topoisomerasa de ARN putante. Participación en el procesamiento de ARN | |||||||
| gyrase inversa
(Archaea) | Elimina (-), pero no (+) supercoils, introduce supercoils positivos | Monomer y Heterodimer | Sí. | |||||
| Topoisomerasa I
()H. sapiens) | Tipo IB | Elimina (+) y (-) supercoils; soporta el movimiento de tenedor durante la replicación y transcripción | Monomer | No | No | SS | 3 ' | ±n |
| Topoisomerasa I
(virus vaccinia) | ||||||||
| Topoisomerasa V
(Archaea) | Tipo IC | Relaja (+) y (-) supercoils. Involucrado en reparación de ADN. | Monomer | No | No | SS | 3 ' | ±n |
| Topoisomerasa II
()E. coli) | Tipo IIA | Genera (-) supercoils (la única topoisomerasa conocida por hacer esto) | Heterotetramer | Sí (Mg2+) | Sí. | DS | 5 ' | ±2 |
| Topoisomerasa IV
()E. coli) | Decatena ADN replicado; relaja (+) supercoils más rápido que (-) | Heterotetramer | ||||||
| Topoisomerasa IIα
()H. sapiens) | Esencial; Unlinks entrelazado hija dúplex en replicación; contribuye a la relajación del ADN durante la transcripción | Homodimer | ||||||
| Topoisomerasa IIβ
()H. sapiens) | Papel en la supresión de la recombinación o la transcripción de apoyo en las neuronas | Homodimer | ||||||
| Topoisomerasa VI
(Archaea) | Tipo IIB | Relaja (+) y (-) supercoils; responsable de descalificar los intermediarios de replicación | Heterotetramer | Sí (Mg2+) | Sí. | DS | 5 ' | ±2 |
Papel de la topoisomerasa en la regulación transcripcional
Al menos una topoisomerasa, la ADN topoisomerasa II beta (topo IIβ), tiene una función reguladora en la transcripción de genes. Las roturas de ADN de doble cadena dependientes de Topo IIβ y los componentes de la maquinaria de reparación de daños en el ADN son importantes para la expresión rápida de genes tempranos inmediatos, así como para la regulación de genes que responden a señales. Topo IIβ, con otras enzimas asociadas, parece ser importante para la liberación de ARN polimerasa pausada en genes largos o altamente transcritos.
Topo IIβ en el inicio de la transcripción
Topo IIβ induce roturas de doble cadena (DSB) de ADN inducidas por estímulo que están limitadas a un corto plazo (de 10 minutos a 2 horas) en las regiones promotoras de genes regulados por señales. Estos DSB permiten una rápida regulación ascendente de la expresión de tales genes sensibles a señales en varios sistemas (ver la Tabla a continuación). Estos genes regulados por señales incluyen genes activados en respuesta a la estimulación con estrógeno, suero, insulina, glucocorticoides (como la dexametasona) y activación de neuronas. Cuando se ha reparado la rotura inducida de la doble hebra del ADN, la transcripción del gen sensible a la señal vuelve a un nivel basal bajo.

Se encontró que Topo IIβ y PARP-1 estaban presentes de forma constitutiva en un nivel moderado cerca del sitio de inicio de la transcripción de un promotor de un gen sensible a la señal. Después de que ocurriera la señal, topo IIβ provocó una ruptura de doble cadena y PARP-1 participó en el reemplazo de la histona H1 por HMGB1/HMGA2, que puede promover la transcripción. Topo IIβ y PARP-1 aumentaron en el sitio de la ruptura de doble cadena y los componentes de la vía de reparación del ADN que se une al extremo no homólogo, incluidos DNA-PKcs, Ku70/Ku80 y ADN ligasa IV ensamblados con topo IIβ y PARP-1. Todo este ensamblaje estaba presente en el ADN enlazador adyacente a un solo nucleosoma en la región promotora de un gen (ver Figura). El nucleosoma estaba cerca del sitio de inicio de la transcripción del gen. Los componentes de la vía de reparación del ADN que se une a los extremos no homólogos fueron esenciales para el cierre de la ruptura de la doble cadena del ADN.
| Ligand o agente activador | Gene(s) evaluado para DSBs | DSB después de la activación inducida por la señal | DSB was required for transcription | DSB location | Proteínas presentes en el DSB | Duración del DSB después de la activación inducida por la señal | Refs |
|---|---|---|---|---|---|---|---|
| Estradiol | pS2 | Sí. | Sí. | promotor | topo IIβ, PARP-1, DNA-PKcs, Ku70, Ku80 | 10 minutos | |
| Insulina | FASN | Sí. | Sí. | promotor | topo IIβ, PARP-1, DNA-PKcs, Ku70, Ku80, protein fosfatase 1 (PP1), P/CAF | 3 horas | |
| Calor de choque o suero | HSPA1B, JUN, FOS, EGR1, MYC | Sí. | Sí. | promotor " POLII pausing site | ADN-PKcs, Ku70, γH2AX, TRIM28 | 30 segundos a 5 minutos | |
| Dexamethasone o Estradiol | PS2, MMTV, PLZF, HSD11B2 | Sí. | Sí. | promotor | topo IIβ, PARP-1, DNA-PKcs, Ku70, Ku80, BRG1 | 15 minutos | |
| KCl o NMDA activación de neuronas corticales primarias cultivadas | FOS, EGR1, NPAS4, NR4A1 | Sí. | Sí. | promotor | topo IIβ, PARP-1, DNA-PKcs, Ku70, Ku80, CTCF | Hasta 2 horas | |
| Acondicionamiento del miedo (evaluado en las neuronas de corteza prefontal del ratón hipocampal y medial) | ■ 200 genes con nuevos DSB y expresión regulada | Sí. | DSBs were correlated with transcription | promotores | no probado | 10 minutos y segundo pico a 30 minutos |
Regulación Topo IIβ de la expresión génica
La ARN polimerasa II con frecuencia tiene un sitio de pausa que se encuentra entre 30 y 60 nucleótidos aguas abajo de el sitio de inicio de la transcripción de un gen. Se cree que la pausa de la ARN polimerasa II en estos sitios y la liberación controlada de la pausa tienen un papel regulador en la transcripción génica. Como señalaron Singh et al., "alrededor del 80 % de los genes altamente expresados en las células HeLa están en pausa". Se producen roturas de doble cadena de ADN inducidas por topo IIβ a muy corto plazo, pero no reselladas inmediatamente, en los sitios de pausa de la ARN polimerasa II, y parecen ser necesarias para la liberación eficiente del estado de pausa y la progresión a la transcripción génica. Para los genes en los que se produce, se cree que la rotura de doble cadena del ADN inducida por TOP2B forma parte del proceso de regulación de la expresión génica.
Contenido relacionado
Pera
Asimina
Célula vegetal