
Oído interno

El oído interno (oído interno, auris interna) es la parte más interna del oído de los vertebrados. En los vertebrados, el oído interno es el principal responsable de la detección y el equilibrio del sonido. En los mamíferos, consiste en el laberinto óseo, una cavidad hueca en el hueso temporal del cráneo con un sistema de pasajes que comprende dos partes funcionales principales:
- El cochlea, dedicado a la audición; convertir patrones de presión del sonido del oído externo en impulsos electroquímicos que se transmiten al cerebro a través del nervio auditivo.
- El sistema vestibular, dedicado al equilibrio
El oído interno se encuentra en todos los vertebrados, con variaciones sustanciales en forma y función. El oído interno está inervado por el octavo nervio craneal en todos los vertebrados.
Estructura

El laberinto se puede dividir por capa o por región.
Laberintos óseos y membranosos
El laberinto óseo, o laberinto óseo, es la red de pasajes con paredes óseas revestidas de periostio. Las tres partes principales del laberinto óseo son el vestíbulo del oído, los canales semicirculares y la cóclea. El laberinto membranoso corre dentro del laberinto óseo y crea tres espacios paralelos llenos de líquido. Los dos exteriores están llenos de perilinfa y el interior de endolinfa.
Sistemas vestibular y coclear
En el oído medio, la energía de las ondas de presión se traduce en vibraciones mecánicas por los tres osículos auditivos. Las ondas de presión mueven la membrana timpánica que a su vez mueve el martillo, el primer hueso del oído medio. El martillo se articula con el yunque que se conecta al estribo. La base del estribo se conecta a la ventana oval, el comienzo del oído interno. Cuando el estribo presiona la ventana oval, hace que la perilinfa, el líquido del oído interno, se mueva. El oído medio sirve así para convertir la energía de las ondas de presión del sonido en una fuerza sobre la perilinfa del oído interno. La ventana oval tiene solo aproximadamente 1/18 del área de la membrana timpánica y, por lo tanto, produce una presión más alta. La cóclea propaga estas señales mecánicas como ondas en el líquido y las membranas y luego las convierte en impulsos nerviosos que se transmiten al cerebro.
El sistema vestibular es la región del oído interno donde convergen los canales semicirculares, cerca de la cóclea. El sistema vestibular trabaja con el sistema visual para mantener los objetos a la vista cuando se mueve la cabeza. Los receptores articulares y musculares también son importantes para mantener el equilibrio. El cerebro recibe, interpreta y procesa la información de todos estos sistemas para crear la sensación de equilibrio.
El sistema vestibular del oído interno es responsable de las sensaciones de equilibrio y movimiento. Utiliza los mismos tipos de fluidos y células de detección (células ciliadas) que utiliza la cóclea y envía información al cerebro sobre la actitud, la rotación y el movimiento lineal de la cabeza. El tipo de movimiento o actitud que detecta una célula ciliada depende de sus estructuras mecánicas asociadas, como el tubo curvo de un canal semicircular o los cristales de carbonato de calcio (otolito) del sáculo y el utrículo.
Desarrollo
El oído interno humano se desarrolla durante la semana 4 del desarrollo embrionario a partir de la placoda auditiva, un engrosamiento del ectodermo que da origen a las neuronas bipolares de los ganglios coclear y vestibular. A medida que la placoda auditiva se invagina hacia el mesodermo embrionario, forma la vesícula auditiva u otoquiste.
La vesícula auditiva dará lugar a los componentes utricular y sacular del laberinto membranoso. Contienen las células ciliadas sensoriales y los otolitos de la mácula del utrículo y del sáculo, respectivamente, que responden a la aceleración lineal ya la fuerza de la gravedad. La división utricular de la vesícula auditiva también responde a la aceleración angular, así como el saco y conducto endolinfático que conecta el sáculo y el utrículo.
A partir de la quinta semana de desarrollo, la vesícula auditiva también da origen al conducto coclear, que contiene el órgano espiral de Corti y la endolinfa que se acumula en el laberinto membranoso. La pared vestibular separará el conducto coclear de la rampa vestibular perilinfática, una cavidad dentro de la cóclea. La membrana basilar separa el conducto coclear de la rampa timpánica, una cavidad dentro del laberinto coclear. La pared lateral del conducto coclear está formada por el ligamento espiral y la estría vascular, que produce la endolinfa. Las células ciliadas se desarrollan a partir de las crestas lateral y medial del conducto coclear, que junto con la membrana tectoria forman el órgano de Corti.
Microanatomía


El canal de Rosenthal o el canal espiral de la cóclea es una sección del laberinto óseo del oído interno que tiene aproximadamente 30 mm de largo y da 2¾ vueltas alrededor del modiolo, el eje central de la cóclea que contiene el ganglio espiral.
Las células del oído interno especializadas incluyen: células ciliadas, células pilares, células de Boettcher, Claudius N#39; células, neuronas del ganglio espiral y Deiters' células (células falángicas).
Las células ciliadas son las células receptoras auditivas primarias y también se conocen como células sensoriales auditivas, células ciliadas acústicas, células auditivas o células de Corti. El órgano de Corti está revestido con una sola fila de células ciliadas internas y tres filas de células ciliadas externas. Las células ciliadas tienen un haz de cabello en la superficie apical de la célula. El haz de cabello consiste en una serie de estereocilios basados en actina. Cada estereocilio se inserta como una raicilla en una densa malla de actina filamentosa conocida como placa cuticular. La interrupción de estos haces da como resultado deficiencias auditivas y defectos del equilibrio.
Las células pilares internas y externas del órgano de Corti sustentan las células ciliadas. Las celdas del pilar externo son únicas porque son celdas independientes que solo están en contacto con las celdas adyacentes en las bases y los vértices. Ambos tipos de células pilares tienen miles de microtúbulos reticulados y filamentos de actina en orientación paralela. Proporcionan acoplamiento mecánico entre la membrana basal y los mecanorreceptores de las células ciliadas.
Las células de Boettcher se encuentran en el órgano de Corti, donde están presentes solo en el giro inferior de la cóclea. Se encuentran en la membrana basilar debajo de Claudius' células y se organizan en filas, cuyo número varía entre especies. Las células se interdigitan entre sí y proyectan microvellosidades en el espacio intercelular. Son células de soporte para las células ciliadas auditivas en el órgano de Corti. Llevan el nombre del patólogo alemán Arthur Böttcher (1831-1889).
Claudio' Las células se encuentran en el órgano de Corti ubicado sobre las filas de células de Boettcher. Al igual que las células de Boettcher, se consideran células de apoyo para las células ciliadas auditivas en el órgano de Corti. Contienen una variedad de canales de agua de acuaporina y parecen estar involucrados en el transporte de iones. También juegan un papel en el sellado de los espacios endolinfáticos. Llevan el nombre del anatomista alemán Friedrich Matthias Claudius (1822-1869).
Deiters' Las células (células falángicas) son un tipo de células neurogliales que se encuentran en el órgano de Corti y están organizadas en una fila de células falángicas internas y tres filas de células falángicas externas. Son las células de soporte del área de células ciliadas dentro de la cóclea. Reciben su nombre del patólogo alemán Otto Deiters (1834-1863) que los describió.
Las celdas de Hensen son celdas columnares altas que están directamente adyacentes a la tercera fila de celdas de Deiters.
La raya de Hensen es la sección de la membrana tectorial sobre la célula ciliada interna.
Los espacios de Nuel se refieren a los espacios llenos de líquido entre las células del pilar externo y las células ciliadas adyacentes y también a los espacios entre las células ciliadas externas.
La membrana de Hardesty es la capa de la tectoria más cercana a la lámina reticular y que recubre la región externa de las células ciliadas.
La membrana de Reissner está compuesta por dos capas de células y separa la escala media de la escala vestibular.
Los dientes de Huschke son las crestas en forma de diente en el limbo espiral que están en contacto con la tectoria y separados por células interdentales.
Suministro de sangre
El laberinto óseo recibe su riego sanguíneo de tres arterias: 1- Rama timpánica anterior (de la arteria maxilar). 2- Rama petrosa (de la arteria meníngea media). 3- Rama estilomastoidea (de la arteria auricular posterior). El laberinto membranoso está irrigado por la arteria laberíntica. El drenaje venoso del oído interno se realiza a través de la vena laberíntica, que desemboca en el seno sigmoideo o seno petroso inferior.
Función
Las neuronas dentro del oído responden a tonos simples y el cerebro sirve para procesar otros sonidos cada vez más complejos. Un adulto promedio generalmente puede detectar sonidos que oscilan entre 20 y 20 000 Hz. La capacidad de detectar sonidos de tonos más altos disminuye en los humanos mayores.
El oído humano ha evolucionado con dos herramientas básicas para codificar ondas sonoras; cada uno está separado para detectar sonidos de alta y baja frecuencia. Georg von Békésy (1899-1972) empleó el uso de un microscopio para examinar la membrana basilar ubicada dentro del oído interno de los cadáveres. Encontró que el movimiento de la membrana basilar se parece al de una onda viajera; cuya forma varía según la frecuencia del tono. En los sonidos de baja frecuencia, la punta (ápice) de la membrana se mueve más, mientras que en los sonidos de alta frecuencia, la base de la membrana se mueve más.
Trastornos
La interferencia o la infección del laberinto puede provocar un síndrome de dolencias llamado laberintitis. Los síntomas de la laberintitis incluyen náuseas, desorientación, vértigo y mareos temporales. La laberintitis puede ser causada por infecciones virales, infecciones bacterianas o bloqueo físico del oído interno.
Otra afección se conoce como enfermedad autoinmune del oído interno (AIED, por sus siglas en inglés). Se caracteriza por hipoacusia neurosensorial bilateral idiopática, rápidamente progresiva. Es un trastorno bastante raro y, al mismo tiempo, la falta de pruebas de diagnóstico adecuadas ha significado que no se puede determinar su incidencia precisa.
Otros animales
Las aves tienen un sistema auditivo similar al de los mamíferos, incluida la cóclea. Los reptiles, los anfibios y los peces no tienen cóclea, pero escuchan con órganos auditivos u órganos vestibulares más simples, que generalmente detectan sonidos de menor frecuencia que la cóclea. La cóclea de las aves también es similar a la de los cocodrilos y consta de un tubo óseo corto y ligeramente curvado dentro del cual se encuentra la membrana basilar con sus estructuras sensoriales.
Sistema coclear
En los reptiles, el sonido se transmite al oído interno a través del hueso del estribo (estribo) del oído medio. Este se presiona contra la ventana oval, una abertura cubierta por una membrana en la superficie del vestíbulo. Desde aquí, las ondas sonoras se conducen a través de un conducto perilinfático corto hasta una segunda abertura, la ventana redonda, que iguala la presión y permite que el fluido incompresible se mueva libremente. En paralelo con el conducto perilinfático hay un conducto ciego separado, el lagena, lleno de endolinfa. La lagena está separada del conducto perilinfático por una membrana basilar y contiene las células ciliadas sensoriales que finalmente traducen las vibraciones del líquido en señales nerviosas. Está unido por un extremo al sáculo.
En la mayoría de los reptiles, el conducto perilinfático y la lagena son relativamente cortos, y las células sensoriales están confinadas a una pequeña papila basilar que se encuentra entre ellos. Sin embargo, en mamíferos, aves y cocodrilos, estas estructuras se vuelven mucho más grandes y algo más complicadas. En aves, cocodrilos y monotremas, los conductos simplemente se extienden, formando juntos un tubo alargado, más o menos recto. El conducto endolinfático se envuelve en un bucle simple alrededor de la lagena, con la membrana basilar a lo largo de un lado. La primera mitad del conducto ahora se denomina rampa vestibular, mientras que la segunda mitad, que incluye la membrana basilar, se denomina rampa timpánica. Como resultado de este aumento de longitud, la membrana basilar y la papila se extienden, y esta última se convierte en el órgano de Corti, mientras que la lagena ahora se denomina conducto coclear. Todas estas estructuras juntas constituyen la cóclea.
En los mamíferos terios, la lagena se extiende aún más, convirtiéndose en una estructura enrollada (cóclea) para acomodar su longitud dentro de la cabeza. El órgano de Corti también tiene una estructura más compleja en los mamíferos que en otros amniotas.
La disposición del oído interno en los anfibios vivos es, en muchos aspectos, similar a la de los reptiles. Sin embargo, a menudo carecen de una papila basilar y, en cambio, tienen un conjunto completamente separado de células sensoriales en el borde superior del sáculo, denominado papilla amphibiorum, que parecen tener la misma función.
Aunque muchos peces son capaces de oír, la lagena es, en el mejor de los casos, un pequeño divertículo del sáculo y parece no tener ningún papel en la sensación del sonido. En cambio, varios grupos de células ciliadas dentro del oído interno pueden ser los responsables; por ejemplo, los peces óseos contienen un grupo sensorial llamado macula negligencia en el utrículo que puede tener esta función. Aunque los peces no tienen oído externo ni medio, el sonido aún puede transmitirse al oído interno a través de los huesos del cráneo o de la vejiga natatoria, partes de las cuales a menudo se encuentran cerca del cuerpo.
Sistema vestibular
En comparación con el sistema coclear, el sistema vestibular varía relativamente poco entre los distintos grupos de vertebrados con mandíbula. La parte central del sistema consta de dos cámaras, el sáculo y el utrículo, cada una de las cuales incluye uno o dos pequeños grupos de células ciliadas sensoriales. Todos los vertebrados con mandíbula también poseen tres canales semicirculares que surgen del utrículo, cada uno con una ampolla que contiene células sensoriales en un extremo.
Un conducto endolinfático va desde el sáculo hasta la cabeza y termina cerca del cerebro. En los peces cartilaginosos, este conducto en realidad se abre en la parte superior de la cabeza, y en algunos teleósteos, es simplemente un final ciego. En todas las demás especies, sin embargo, termina en un saco endolinfático. En muchos reptiles, peces y anfibios, este saco puede alcanzar un tamaño considerable. En los anfibios, los sacos de ambos lados pueden fusionarse en una sola estructura, que a menudo se extiende a lo largo del cuerpo, paralela al canal espinal.
Las lampreas y los mixinos primitivos, sin embargo, tienen un sistema más simple. El oído interno en estas especies consta de una sola cámara vestibular, aunque en las lampreas está asociada con una serie de sacos revestidos por cilios. Las lampreas tienen solo dos canales semicirculares, sin el canal horizontal, mientras que los mixinos tienen solo un canal vertical.
Equilibrio
El oído interno es el principal responsable del equilibrio, el equilibrio y la orientación en el espacio tridimensional. El oído interno puede detectar tanto el equilibrio estático como el dinámico. Tres conductos semicirculares y dos cámaras, que contienen el sáculo y el utrículo, permiten al cuerpo detectar cualquier desviación del equilibrio. La macula sacculi detecta la aceleración vertical mientras que la macula utriculi es responsable de la aceleración horizontal. Estas estructuras microscópicas poseen estereocilios y un cinocilio que se encuentran dentro de la membrana otolítica gelatinosa. La membrana se carga aún más con otolitos. El movimiento de los estereocilios y el kinocilio permite que las células ciliadas del sáculo y el utrículo detecten el movimiento. Los conductos semicirculares son los encargados de detectar el movimiento de rotación.
Imágenes adicionales
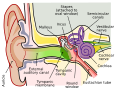 Anatomía del oído humano.Brown es el oído exterior.Rojo es oído medio.El púrpura es oído interno.
Anatomía del oído humano.Brown es el oído exterior.Rojo es oído medio.El púrpura es oído interno.
Laberinto de oído

Oreja interior

Hueso temporal
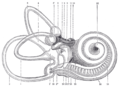
El laberinto membranoso humano derecho, removido de su recinto bony y visto desde el aspecto antero-lateral





Contenido relacionado
Síndrome de Tourette
Disfunción eréctil
Corteza entorrinal