
Moa
Moa (orden Dinornithiformes) son un grupo extinto de aves no voladoras anteriormente endémicas de Nueva Zelanda. Había nueve especies (en seis géneros). Las dos especies más grandes, Dinornis robustus y Dinornis novaezelandiae, alcanzaban unos 3,6 metros (12 pies) de altura con el cuello extendido y pesaban unos 230 kilogramos (510 lb), mientras que la el más pequeño, el moa arbustivo (Anomalopteryx didiformis), tenía aproximadamente el tamaño de un pavo. Las estimaciones de la población moa cuando los polinesios se establecieron en Nueva Zelanda alrededor de 1300 varían entre 58.000 y aproximadamente 2,5 millones.
Las moa se colocan tradicionalmente en el grupo de las rátidas. Sin embargo, sus parientes más cercanos han sido encontrados por estudios genéticos como el tinamú sudamericano volador, una vez considerado como un grupo hermano de las ratites. Las nueve especies de moa eran las únicas aves sin alas, que carecían incluso de las alas vestigiales que tienen todas las demás ratites. Eran los animales terrestres más grandes y los herbívoros dominantes en los bosques, matorrales y ecosistemas subalpinos de Nueva Zelanda hasta la llegada de los maoríes, y solo eran cazados por el águila de Haast. La extinción de Moa ocurrió dentro de los 100 años del asentamiento humano de Nueva Zelanda, principalmente debido a la caza excesiva.
Etimología
La palabra moa es un término polinesio para las aves domésticas. El nombre no era de uso común entre los maoríes en el momento del contacto con los europeos, probablemente porque el ave que describía se había extinguido durante algún tiempo y las historias tradicionales al respecto eran raras. El registro más antiguo del nombre fue de los misioneros William Williams y William Colenso en enero de 1838; Colenso especuló que las aves podrían haberse parecido a aves gigantes. En 1912, el jefe maorí Urupeni Pūhara afirmó que el nombre tradicional de los moa era "te kura" (el pájaro rojo).
Descripción

Los esqueletos de moa se reconstruían tradicionalmente en posición vertical para crear una altura impresionante, pero el análisis de sus articulaciones vertebrales indica que probablemente llevaban la cabeza hacia adelante, a la manera de un kiwi. La columna vertebral estaba unida a la parte posterior de la cabeza en lugar de la base, lo que indica la alineación horizontal. Esto les habría permitido pastar en la vegetación baja, mientras podían levantar la cabeza y buscar árboles cuando era necesario. Esto ha resultado en una reconsideración de la altura de los moa más grandes. Sin embargo, el arte rupestre maorí representa moa o pájaros parecidos a moa (probablemente gansos o adzebills) con el cuello erguido, lo que indica que los moa eran más que capaces de asumir ambas posturas del cuello.
No sobreviven registros de los sonidos que hacían los moa, aunque se puede obtener una idea de sus llamadas a partir de la evidencia fósil. La tráquea de moa estaba sostenida por muchos pequeños anillos de hueso conocidos como anillos traqueales. La excavación de estos anillos de esqueletos articulados ha demostrado que al menos dos géneros de moa (Euryapteryx y Emeus) exhibieron elongación traqueal, es decir, su tráquea era de hasta 1 m (3 ft) de largo y formó un gran bucle dentro de la cavidad del cuerpo. Son las únicas ratites conocidas que exhiben esta característica, que también está presente en varios otros grupos de aves, incluidos los cisnes, las grullas y las pintadas. La característica está asociada con vocalizaciones resonantes profundas que pueden viajar largas distancias.
Relaciones evolutivas

Los parientes más cercanos del moa son pequeñas aves terrestres sudamericanas llamadas tinamú, que pueden volar. Anteriormente, se pensaba que el kiwi, el emú australiano y el casuario estaban más estrechamente relacionados con el moa.
Aunque se describieron docenas de especies a fines del siglo XIX y principios del XX, muchas se basaron en esqueletos parciales y resultaron ser sinónimos. Actualmente, 11 especies están reconocidas formalmente, aunque estudios recientes que utilizan ADN antiguo recuperado de huesos en colecciones de museos sugieren que existen distintos linajes dentro de algunos de estos. Un factor que ha causado mucha confusión en la taxonomía de los moa es la variación intraespecífica del tamaño de los huesos, entre períodos glaciales e interglaciales (ver la regla de Bergmann y la regla de Allen), así como el dimorfismo sexual que es evidente en varias especies. Dinornis parece haber tenido el dimorfismo sexual más pronunciado, siendo las hembras hasta un 150 % más altas y un 280 % más pesadas que los machos, tanto más grandes que fueron clasificadas como especies separadas hasta 2003. A 2009 estudio mostró que Euryapteryx curtus y E. gravis eran sinónimos. Un estudio de 2010 explicó las diferencias de tamaño entre ellos como dimorfismo sexual. En cambio, un estudio morfológico de 2012 los interpretó como subespecies.
Los análisis de ADN antiguo han determinado que varios linajes evolutivos crípticos ocurrieron en varios géneros de moa. Estos pueden eventualmente clasificarse como especies o subespecies; Megalapteryx benhami (Archey) es sinónimo de M. didinus (Owen) porque los huesos de ambos comparten todos los caracteres esenciales. Las diferencias de tamaño pueden explicarse por un cline norte-sur combinado con una variación temporal, de modo que los especímenes eran más grandes durante el período glacial de Otiran (la última edad de hielo en Nueva Zelanda). Se conoce una variación de tamaño temporal similar para Pachyornis mappini de la Isla Norte. Algunas de las otras variaciones de tamaño de las especies de moa probablemente puedan explicarse por factores geográficos y temporales similares.
Los primeros restos de moa proceden de la fauna del Mioceno Saint Bathans. Conocidos por múltiples cáscaras de huevo y elementos de las patas traseras, estos representan al menos dos especies que ya tienen un tamaño bastante grande.
Clasificación
Taxonomía

_601344.jpg)
Los géneros y especies actualmente reconocidos son:
- Orden †Dinornithiformes (Gadow 1893) Ridgway 1901 [Dinornithes Gadow 1893; Immanes Newton 1884] (moa)
- Familia Dinornithidae Owen 1843 [Palapteryginae Bonaparte 1854; Palapterygidae Haast 1874; Dinornithnideae Stejneger 1884] (giant moa)
- Genus Dinornis
- North Island gigante moa, Dinornis novaezealandiae (Isla Norte, Nueva Zelandia)
- Musa gigante de la Isla del Sur, Dinornis robustus (South Island, Nueva Zelandia)
- Genus Dinornis
- Familia Emeidae (Bonaparte 1854) [Emeinae] Bonaparte 1854; Anomalopterygidae Oliver 1930; Anomalapteryginae Archey 1941]
- Genus Anomalopteryx
- Bush Moa, Anomalopteryx didiformis (Isla Norte y Sur, Nueva Zelandia)
- Genus Emeus
- Moa oriental, Emeus crasus (South Island, Nueva Zelandia)
- Genus Euryapteryx
- Moa de gran alcance, Euryapteryx curtus (Isla Norte y Sur, Nueva Zelandia)
- Genus Pachyornis
- Moa de pies pesados, Pachyornis elephantopus (South Island, Nueva Zelandia)
- Mantell's moa, Pachyornis geranoides (Isla Norte, Nueva Zelandia)
- Moa crecida, Pachyornis australis (South Island, Nueva Zelandia)
- Genus Anomalopteryx
- Familia Megalapterygidae
- Genus Megalapteryx
- Upland moa, Megalapteryx didinus (South Island, Nueva Zelandia)
- Genus Megalapteryx
- Familia Dinornithidae Owen 1843 [Palapteryginae Bonaparte 1854; Palapterygidae Haast 1874; Dinornithnideae Stejneger 1884] (giant moa)
Dos especies sin nombre de la fauna de Saint Bathans.
Filogenia
Debido a que los moa son un grupo de aves no voladoras sin vestigios de huesos en las alas, se han planteado dudas sobre cómo llegaron a Nueva Zelanda y de dónde. Existen muchas teorías sobre la llegada y la radiación de los moas a Nueva Zelanda, pero la teoría más reciente sugiere que llegaron a Nueva Zelanda hace unos 60 millones de años (Mya) y se separaron de la región "basal" (ver más abajo) especies de moa, Megalapteryx, alrededor de 5,8 Mya en lugar de la división de 18,5 Mya sugerida por Baker et al. (2005). Esto no significa necesariamente que no hubo especiación entre la llegada 60 Mya y la división basal 5,8 Mya, pero falta el registro fósil y lo más probable es que existieran los primeros linajes moa, pero se extinguieron antes de la división basal 5,8 Mya. La presencia de especies del Mioceno ciertamente sugiere que la diversificación de moa comenzó antes de la división entre Megalapteryx y los otros taxones.
El evento máximo de ahogamiento del Oligoceno, que ocurrió hace aproximadamente 22 millones de años, cuando solo el 18 % de la actual Nueva Zelanda estaba sobre el nivel del mar, es muy importante en la radiación moa. Debido a que la división del moa basal ocurrió tan recientemente (5,8 millones de años), se argumentó que los ancestros de los linajes de moa del Cuaternario no podrían haber estado presentes en los remanentes de la Isla Norte y del Sur durante el ahogamiento del Oligoceno. Esto no implica que los moa estuvieran previamente ausentes de la Isla Norte, sino que solo sobrevivieron los de la Isla Sur, porque solo la Isla Sur estaba sobre el nivel del mar. Bunce et al. (2009) argumentaron que los ancestros moa sobrevivieron en la Isla Sur y luego recolonizaron la Isla Norte alrededor de 2 Ma más tarde, cuando las dos islas se volvieron a unir después de 30 Ma de separación. La presencia de moa del Mioceno en la fauna de Saint Bathans parece sugerir que estas aves aumentaron de tamaño poco después del evento de ahogamiento del Oligoceno, si es que se vieron afectadas por él.
Bunce et al. también concluyó que la estructura altamente compleja del linaje moa fue causada por la formación de los Alpes del Sur alrededor de 6 millones de años y la fragmentación del hábitat en ambas islas como resultado de los ciclos glaciales del Pleistoceno, el vulcanismo y los cambios en el paisaje. El cladograma a continuación es una filogenia de Palaeognathae generada por Mitchell (2014) con algunos nombres de clados después de Yuri et al. (2013). Proporciona la posición de la moa (Dinornithiformes) dentro del contexto más amplio de la "mandíbula antigua" (Palaeognathae) aves:
| Palaeognathae |
| ||||||||||||||||||||||||||||||||||||||||||
El cladograma a continuación brinda una filogenia más detallada, a nivel de especie, de la rama moa (Dinornithiformes) del "mandíbula antigua" aves (Palaeognathae) que se muestran arriba:
| †Dinornithiformes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Distribución y hábitat
Los análisis de conjuntos de huesos de moa fósiles han proporcionado datos detallados sobre las preferencias de hábitat de las especies de moa individuales y han revelado faunas de moa regionales distintivas:
Isla Sur

Las dos faunas principales identificadas en la Isla Sur incluyen:
- La fauna de la alta maraña de la costa oesteNothofagus) bosques que incluyeron Anomalopteryx didiformis (bush moa) y Dinornis robustus (Moa gigante de la Isla del Sur) y
- La fauna del bosque de sombras de lluvias y arbustos al este de los Alpes del Sur que incluía Pachyornis elephantopus (Moa de patas pesadas), Euryapteryx gravis, Emeus crasus, y Dinornis robustus.
Una 'fauna subalpina' podría incluir la generalizada D. robustus, y las otras dos especies de moa que existían en la Isla Sur:
- Pachyornis australis, las especies de moa más raras, las únicas especies de moa que aún no se encuentran en los middens maoríes. Sus huesos se han encontrado en cuevas en el noroeste de Nelson y los distritos de Karamea (como la Cueva de la Colina Honeycomb), y algunos sitios alrededor del distrito de Wānaka.
- Megalapteryx didinus, más generalizado, llamado "moa de tierra" porque sus huesos se encuentran comúnmente en la zona subalpina. Sin embargo, también ocurrió a nivel del mar, donde existían terrenos empinados y rocosos adecuados (como Punakaiki en la costa oeste y Otago Central). Sus distribuciones en zonas costeras han sido bastante poco claras, pero estuvieron presentes al menos en varios lugares como Kaikōura, Otago Peninsula y Karitane.

Isla Norte
Se sabe mucho menos sobre las paleofaunas de la Isla Norte, debido a la escasez de yacimientos de fósiles en comparación con la Isla Sur, pero el patrón básico de las relaciones moa-hábitat era el mismo. La Isla Sur y la Isla Norte compartían algunas especies de moa (Euryapteryx gravis, Anomalopteryx didiformis), pero la mayoría eran exclusivas de una isla, lo que refleja la divergencia durante varios miles de años desde el nivel inferior del mar. nivel en la edad de hielo había hecho un puente de tierra a través del estrecho de Cook.
En la Isla Norte, Dinornis novaezealandiae y Anomalopteryx didiformis dominaron el hábitat del bosque de alta precipitación, un patrón similar al de la Isla Sur. Las otras especies de moa presentes en la Isla Norte (Euryapteryx gravis, E. curtus y Pachyornis geranoides) tendían a habitar hábitats de bosques y matorrales más secos. P. geranoides ocurrieron en toda la Isla Norte. Las distribuciones de E. gravis y E. curtus eran casi mutuamente excluyentes, ya que el primero solo se encontró en sitios costeros alrededor de la mitad sur de la Isla Norte.
Comportamiento y ecología

Se han encontrado alrededor de ocho huellas de moa, con huellas de moa fosilizadas en sedimentos fluviales, en la Isla Norte, incluidos Waikanae Creek (1872), Napier (1887), Manawatū River (1895), Marton (1896), Palmerston North (1911) (ver fotografía a la izquierda), río Rangitīkei (1939) y bajo el agua en el lago Taupō (1973). El análisis del espaciado de estas huellas indica velocidades de caminata entre 3 y 5 km/h (1,75–3 mph).
Dieta

Su dieta se ha deducido a partir del contenido fosilizado de sus mollejas y coprolitos, así como indirectamente a través del análisis morfológico del cráneo y el pico, y el análisis de isótopos estables de sus huesos. Moa se alimentaba de una variedad de especies de plantas y partes de plantas, incluidas ramitas fibrosas y hojas extraídas de árboles y arbustos bajos. El pico de Pachyornis elephantopus era análogo a un par de tijeras de podar y podía cortar las hojas fibrosas del lino de Nueva Zelanda (Phormium tenax) y ramitas de hasta al menos 8 mm de diámetro. diámetro.
Moa llenó el nicho ecológico ocupado en otros países por grandes mamíferos ramoneadores como antílopes y llamas. Algunos biólogos sostienen que varias especies de plantas evolucionaron para evitar el ramoneo de moa. Plantas divaracantes como Pennantia corymbosa (la kaikōmako), que tiene hojas pequeñas y una densa malla de ramas, y Pseudopanax crassifolius (la horoeka o lancewood), que tiene una dura hojas, son posibles ejemplos de plantas que evolucionaron de esa manera.
Al igual que muchas otras aves, el moa tragaba piedras de molleja (gastrolitos), que quedaban retenidas en sus mollejas musculares, proporcionando una acción trituradora que les permitía comer material vegetal grueso. Estas piedras eran comúnmente guijarros de cuarzo redondeados y lisos, pero se han encontrado piedras de más de 110 milímetros (4 pulgadas) de largo entre los contenidos conservados de mollejas de moa. Las mollejas de Dinornis a menudo podían contener varios kilogramos de piedras. Moa probablemente ejerció cierta selectividad en la elección de las piedras de molleja y eligió los guijarros más duros.
Reproducción
Las parejas de especies de moa descritas como Euryapteryx curtus / E. exilis, Emeus huttonii / E. crassus, y Pachyornis septentrionalis / P. mappini se ha sugerido durante mucho tiempo que constituyen machos y hembras, respectivamente. Esto ha sido confirmado por el análisis de marcadores genéticos específicos del sexo del ADN extraído del material óseo.
Por ejemplo, antes de 2003, se reconocían tres especies de Dinornis: moa gigante de la Isla Sur (D. robustus), moa gigante de la Isla Norte (D. novaezealandiae), y moa esbelta (D. struthioides). Sin embargo, el ADN mostró que todos los D. struthioides eran machos, y todos D. robustus eran hembras. Por lo tanto, las tres especies de Dinornis se reclasificaron como dos especies, cada una de las cuales ocurría anteriormente en la Isla Norte de Nueva Zelanda (D. novaezealandiae) y en la Isla Sur (D. robustus); D. robustus sin embargo, comprende tres linajes genéticos distintos y eventualmente puede clasificarse como muchas especies, como se discutió anteriormente.
El examen de los anillos de crecimiento en el hueso cortical de moa ha revelado que estas aves fueron seleccionadas con K, al igual que muchas otras aves grandes endémicas de Nueva Zelanda. Se caracterizan por tener una baja fecundidad y un largo período de maduración, tardando unos 10 años en alcanzar el tamaño adulto. Las especies grandes de Dinornis tardaron tanto en alcanzar el tamaño adulto como las especies pequeñas de moa y, como resultado, tuvieron un rápido crecimiento esquelético durante sus años juveniles.
No se ha encontrado ninguna evidencia que sugiera que los moa fueran nidos coloniales. La anidación de Moa a menudo se infiere de las acumulaciones de fragmentos de cáscara de huevo en cuevas y refugios rocosos, existe poca evidencia de los nidos en sí. Las excavaciones de refugios rocosos en el este de la Isla Norte durante la década de 1940 encontraron nidos de moa, que se describieron como "pequeñas depresiones obviamente talladas en la piedra pómez suave y seca". El material de anidación de moa también se ha recuperado de refugios rocosos en la región central de Otago en la Isla Sur, donde el clima seco ha preservado el material vegetal utilizado para construir la plataforma de anidación (incluidas ramitas cortadas por picos de moa). Las semillas y el polen dentro de los coprolitos de moa que se encuentran entre el material de anidación proporcionan evidencia de que la temporada de anidación fue desde fines de la primavera hasta el verano.
A menudo se encuentran fragmentos de cáscara de huevo de moa en sitios arqueológicos y dunas de arena alrededor de la costa de Nueva Zelanda. Treinta y seis huevos de moa enteros existen en colecciones de museos y varían mucho en tamaño (de 120 a 240 milímetros (4,7 a 9,4 pulgadas) de largo y de 91 a 178 milímetros (3,6 a 7,0 pulgadas) de ancho). La superficie exterior de la cáscara de huevo de moa se caracteriza por pequeños poros en forma de hendidura. Los huevos de la mayoría de las especies de moa eran blancos, aunque los del moa de las tierras altas (Megalapteryx didinus) eran de color verde azulado.
Un estudio de 2010 realizado por Huynen et al. descubrió que los huevos de ciertas especies eran frágiles, solo alrededor de un milímetro de grosor de la cáscara: "Inesperadamente, también se demostró que varios huevos de cáscara delgada pertenecían al moa más pesado del género Dinornis, Euryapteryx y Emeus, lo que los convierte, según nuestro conocimiento, en los más frágiles de todos los huevos de aves medidos hasta la fecha. Además, el ADN específico del sexo recuperado de las superficies exteriores de las cáscaras de huevo pertenecientes a especies de Dinornis y Euryapteryx sugiere que estos huevos muy delgados probablemente fueron incubados por los machos más ligeros.. La naturaleza delgada de las cáscaras de huevo de estas especies más grandes de moa, incluso si fueron incubadas por el macho, sugiere que la rotura de huevos en estas especies habría sido común si se hubiera utilizado el método de contacto típico de la incubación de huevos de aves. A pesar de la extinción del ave, el alto rendimiento de ADN disponible de los huevos fosilizados recuperados ha permitido secuenciar el genoma del moa.

El esqueleto de moa hembra de tierra firme con huevo en posición unlaid dentro de la cavidad pélvica en el Museo Otago
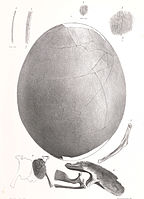
Un óvulo y fragmentos embriones de Emeus crasus

Restauración de un moa continental



Bosques prehumanos
Los estudios de vegetación seca acumulada en el período prehumano del Holoceno medio tardío sugieren un ecosistema de bosque bajo de Sophora microphylla o Kōwai en Central Otago que fue utilizado y quizás mantenido por moa, tanto para material de anidación y comida. Ni los bosques ni los moa existían cuando los colonos europeos llegaron a la zona en la década de 1850.
Relación con los humanos
Extinción

Antes de la llegada de los colonos humanos, el único depredador del moa era el enorme águila de Haast. Nueva Zelanda había estado aislada durante 80 millones de años y tenía pocos depredadores antes de la llegada de los humanos, lo que significa que sus ecosistemas no solo eran extremadamente vulnerables a la perturbación de especies externas, sino que las especies nativas estaban mal equipadas para hacer frente a los depredadores humanos.
Los polinesios llegaron en algún momento antes de 1300, y todos los géneros de moa pronto fueron llevados a la extinción por la caza y, en menor medida, por la reducción del hábitat debido a la tala de bosques. Para 1445, todos los moa se habían extinguido, junto con el águila de Haast, que dependía de ellos para su alimentación. Investigaciones recientes que utilizan la datación por carbono 14 de basureros sugieren fuertemente que los eventos que llevaron a la extinción duraron menos de cien años, en lugar de un período de explotación que duró varios cientos de años, como se había planteado anteriormente.
Algunos autores han especulado que algunos Megalapteryx didinus pueden haber persistido en rincones remotos de Nueva Zelanda hasta los siglos XVIII e incluso XIX, pero esta opinión no es ampliamente aceptada. Algunos cazadores maoríes afirmaron estar persiguiendo al moa hasta la década de 1770; sin embargo, estos relatos posiblemente no se refirieron tanto a la caza de aves reales como a un ritual ahora perdido entre los habitantes de las islas del sur. Los balleneros y cazadores de focas recordaron haber visto aves monstruosas a lo largo de la costa de la Isla Sur, y en la década de 1820, un hombre llamado George Pauley hizo una afirmación no verificada de haber visto un moa en la región de Otago en Nueva Zelanda.
Una expedición en la década de 1850 bajo el mando del teniente A. Impey informó sobre dos pájaros parecidos a emú en una colina en la Isla Sur; una historia de 1861 del Nelson Examiner hablaba de huellas de tres dedos que medían 36 cm (14 pulgadas) entre Tākaka y Riwaka que fueron encontradas por un grupo de inspección; y finalmente en 1878, el Otago Witness publicó un relato adicional de un granjero y su pastor. Una mujer de 80 años, Alice McKenzie, afirmó en 1959 que había visto un moa en los arbustos de Fiordland en 1887 y nuevamente en una playa de Fiordland cuando tenía 17 años. Afirmó que su hermano también había visto un moa en otra ocasión.
Restos sobrevivientes

Joel Polack, un comerciante que vivió en la costa este de la Isla Norte entre 1834 y 1837, registró en 1838 que le habían mostrado "varias osificaciones fósiles grandes" encontrado cerca del monte Hikurangi. Estaba seguro de que se trataba de los huesos de una especie de emú o avestruz, señalando que "los nativos agregan que en tiempos pasados recibieron las tradiciones de que habían existido aves muy grandes, pero la escasez de alimentos para animales, así como como el método fácil de atraparlos, ha provocado su exterminio". Polack señaló además que había recibido informes de los maoríes de que una "especie de Struthio" todavía existía en partes remotas de la Isla Sur.
Dieffenbach también hace referencia a un fósil de la zona cercana al monte Hikurangi, y supone que pertenece a "un pájaro, ahora extinto, llamado Moa (o Movie) por los nativos". 'Película' es el primer nombre transcrito para el ave. En 1839, John W. Harris, un comerciante de lino de Poverty Bay que era un entusiasta de la historia natural, recibió un trozo de hueso inusual de un maorí que lo había encontrado en la orilla de un río. Le mostró el fragmento de hueso de 15 cm (6 pulgadas) a su tío, John Rule, un cirujano de Sydney, quien se lo envió a Richard Owen, quien en ese momento trabajaba en el Museo Hunterian en el Royal College of Surgeons en Londres.

Owen pensó en el fragmento durante casi cuatro años. Estableció que era parte del fémur de un animal grande, pero era inusualmente ligero y con forma de panal. Owen anunció a una comunidad científica escéptica y al mundo que se trataba de un pájaro gigante extinto como un avestruz, y lo llamó Dinornis. Su deducción fue ridiculizada en algunos sectores, pero se demostró que era correcta con los posteriores descubrimientos de cantidades considerables de huesos de moa en todo el país, suficientes para reconstruir los esqueletos de las aves.
En julio de 2004, el Museo de Historia Natural de Londres exhibió el fragmento de hueso de moa que Owen había examinado por primera vez, para celebrar los 200 años desde su nacimiento y en memoria de Owen como fundador del museo.

Desde el descubrimiento de los primeros huesos de moa a fines de la década de 1830, se han encontrado miles más. Ocurren en una variedad de depósitos sedimentarios del Cuaternario tardío y del Holoceno, pero son más comunes en tres tipos principales de sitios: cuevas, dunas y pantanos.

Los huesos se encuentran comúnmente en cuevas o tomo (la palabra maorí para dolina o sumidero, que a menudo se usa para referirse a trampas o pozos verticales de cuevas). Las dos formas principales en que los huesos de moa se depositaron en tales sitios fueron las aves que ingresaron a la cueva para anidar o escapar del mal tiempo y luego murieron en la cueva y las aves que cayeron en un pozo vertical y no pudieron escapar. Se han encontrado huesos de moa (y huesos de otras aves extintas) en cuevas por toda Nueva Zelanda, especialmente en las áreas de piedra caliza/mármol del noroeste de Nelson, Karamea, Waitomo y Te Anau.
Los huesos de moa y los fragmentos de cáscara de huevo a veces se encuentran en las dunas de arena costeras activas, donde pueden erosionarse desde los paleosuelos y concentrarse en 'erupciones' entre cordones de dunas. Muchos de estos huesos de moa son anteriores a los asentamientos humanos, aunque algunos se originan en basureros maoríes, que con frecuencia se encuentran en dunas cerca de puertos y desembocaduras de ríos (por ejemplo, los grandes sitios de cazadores de moa en Shag River, Otago y Wairau Bar, Marlborough).
Se han encontrado huesos de moa densamente entremezclados en pantanos de Nueva Zelanda. El ejemplo más conocido está en Pyramid Valley en el norte de Canterbury, donde se han excavado huesos de al menos 183 moa individuales, en su mayoría por Roger Duff del Museo de Canterbury. Se han propuesto muchas explicaciones para explicar cómo se formaron estos depósitos, que van desde aguas de manantial venenosas hasta inundaciones e incendios forestales. Sin embargo, la explicación actualmente aceptada es que los huesos se acumularon lentamente durante miles de años, de aves que ingresaron a los pantanos para alimentarse y quedaron atrapadas en el sedimento blando.
Muchos museos de Nueva Zelanda e internacionales tienen colecciones de huesos de moa. Museo Conmemorativo de la Guerra de Auckland: Tāmaki Paenga Hira tiene una colección importante, y en 2018 se tomaron imágenes de varios esqueletos de moa y se escanearon en 3D para que las colecciones sean más accesibles. También hay una colección importante en el Museo Otago en Dunedin.
Plumas y tejidos blandos

Se han encontrado varios ejemplos notables de restos de moa que exhiben tejidos blandos (músculos, piel, plumas), que se preservaron a través de la desecación cuando el ave murió en un sitio naturalmente seco (por ejemplo, una cueva con una brisa seca constante que sopla a traves de). La mayoría de estos especímenes se han encontrado en la región semiárida de Central Otago, la parte más seca de Nueva Zelanda. Éstos incluyen:
- Musculo seco en los huesos de una mujer Dinornis robustus encontrado en Tiger Hill en el valle del río Manuherikia por mineros de oro en 1864 (actualmente ocupado por el Museo Yorkshire)
- Varios huesos Emeus crasus con músculo unido, y una hilera de vértebras del cuello con músculo, piel y plumas recolectadas de la cueva de Earnscleugh cerca de la ciudad de Alexandra en 1870 (actualmente sostenida por el Museo Otago)
- Un pie articulado de un macho D. giganteus con almohadillas de piel y pie conservadas, encontradas en un grieta en la cordillera de Knobby en 1874 (actualmente sostenida por el Museo Otago)
- El tipo de muestra Megalapteryx didinus encontrado cerca de Queenstown en 1878 (actualmente realizado por Natural History Museum, Londres; ver la fotografía de pie en esta página)
- La pierna inferior Pachyornis elephantopus, con piel y músculo, de la cordillera Héctor en 1884; (actualmente sostenida por el Departamento de Zoología de la Universidad de Cambridge)
- La pierna llena de plumas M. didinus de Old Man Range en 1894 (actualmente organizado por Otago Museum)
- La cabeza de un M. didinus encontrado cerca de Cromwell en algún momento antes de 1949 (actualmente celebrado por el Museo de Nueva Zelanda).
Se conocen dos especímenes de fuera de la región de Central Otago:
- Un pie completo M. didinus encontrado en una cueva en el Monte Owen cerca de Nelson en los años 80 (actualmente celebrada por el Museo de Nueva Zelanda)
- Un esqueleto Anomalopteryx didiformis con bases musculares, de piel y plumas recogidas de una cueva cerca de Te Anau en 1980.

Además de estos especímenes, se recolectaron plumas de moa sueltas de cuevas y refugios rocosos en el sur de la Isla Sur y, con base en estos restos, se logró una idea del plumaje del moa. La pierna preservada de M. didinus de Old Man Range revela que esta especie estaba emplumada hasta el pie. Es probable que esto haya sido una adaptación a vivir en entornos nevados a gran altitud, y también se ve en el ñandú de Darwin, que vive en un hábitat nevado estacional similar.
Las plumas de moa miden hasta 23 cm (9 pulgadas) de largo y se ha informado de una variedad de colores, que incluyen marrón rojizo, blanco, amarillento y morado. También se han encontrado plumas oscuras con puntas blancas o cremosas, lo que indica que algunas especies de moa pueden haber tenido un plumaje con apariencia moteada.
Afirmaciones de supervivencia

Se ha especulado de vez en cuando, al menos desde finales del siglo XIX y tan recientemente como 1993 y 2008, sobre la posibilidad de que todavía existan algunos moa, en particular en las tierras salvajes de South Westland y Fiordland. El informe de 1993 inicialmente interesó al Departamento de Conservación, pero el animal en una fotografía borrosa fue identificado como un ciervo rojo. Los criptozoólogos continúan buscándolos, pero sus afirmaciones y la evidencia de respaldo (como las supuestas huellas) han ganado poca atención por parte de los expertos y son pseudocientíficas. En 1880, Alice Mackenzie tuvo un encuentro con un pájaro grande que creía que era un takahē, pero cuando fue redescubierto en la década de 1940, y Mackenzie vio lo que parecía, supo que había visto algo más.
El redescubrimiento del takahē en 1948 después de que no se había visto ninguno desde 1898 mostró que las aves raras pueden existir sin ser descubiertas durante mucho tiempo. Sin embargo, el takahē es un pájaro mucho más pequeño que el moa, y fue redescubierto después de que se identificaron sus huellas; sin embargo, nunca se ha encontrado evidencia confiable de huellas de moa, y los expertos aún sostienen que la supervivencia del moa es extremadamente improbable, ya que tendrían que estar viviendo desapercibido durante más de 500 años en una región visitada a menudo por cazadores y excursionistas.
Reactivación potencial
La criatura se ha mencionado con frecuencia como candidata potencial para revivir mediante la clonación. Su estatus icónico, junto con el hecho de que solo se extinguió hace unos cientos de años y que existen cantidades sustanciales de restos de moa, significa que a menudo se incluye junto a criaturas como el dodo como los principales candidatos para la extinción. El genetista japonés Ankoh Yasuyuki Shirota ha llevado a cabo un trabajo preliminar relacionado con la extracción de ADN.
El interés en el potencial de reavivamiento de los moa aumentó aún más a mediados de 2014 cuando el miembro del Parlamento de Nueva Zelanda, Trevor Mallard, sugirió que traer de vuelta algunas especies más pequeñas de moa dentro de 50 años era una idea viable. La idea fue ridiculizada por muchos, pero obtuvo el apoyo de algunos expertos en historia natural.
En literatura y cultura

Heinrich Harder retrató a un moa siendo cazado por maoríes en el clásico alemán de colección de cartas sobre animales prehistóricos y extintos, "Tiere der Urwelt", a principios del siglo XX.
El poema de Allen Curnow, "El esqueleto del Gran Moa en el Museo de Canterbury, Christchurch" fue publicado en 1943.
Contenido relacionado
Narciso (planta)
Rododendro
Rosales