
Microscopio de fluorescencia

A Microscopio de fluorescencia es un microscopio óptico que utiliza fluorescencia en vez de dispersar, reflexionar y atenuación o absorción, para estudiar las propiedades de las sustancias orgánicas o inorgánicas. "Micróscopo de fluorescencia" se refiere a cualquier microscopio que use fluorescencia para generar una imagen, ya sea una simple configuración como un microscopio epifluorescencia o un diseño más complicado como un microscopio confocal, que utiliza la sección óptica para obtener una mejor resolución de la imagen de fluorescencia.
Principio
La muestra se ilumina con luz de una longitud de onda (o longitudes de onda) específica que es absorbida por los fluoróforos, lo que hace que emitan luz de longitudes de onda más largas (es decir, de un color diferente al de la luz absorbida). La luz de iluminación se separa de la fluorescencia emitida, mucho más débil, mediante el uso de un filtro de emisión espectral. Los componentes típicos de un microscopio de fluorescencia son una fuente de luz (la lámpara de arco de xenón o la lámpara de vapor de mercurio son comunes; las formas más avanzadas son los LED y láseres de alta potencia), el filtro de excitación, el espejo dicroico (o divisor de haz dicroico) y el sistema de emisión. filtro (ver figura a continuación). Los filtros y el divisor de haz dicroico se eligen para que coincidan con las características espectrales de excitación y emisión del fluoróforo utilizado para marcar la muestra. De esta manera, se obtienen imágenes de la distribución de un solo fluoróforo (color) a la vez. Se deben componer imágenes multicolores de varios tipos de fluoróforos combinando varias imágenes de un solo color.
La mayoría de los microscopios de fluorescencia que se utilizan son microscopios de epifluorescencia, donde la excitación del fluoróforo y la detección de la fluorescencia se realizan a través del mismo camino de luz (es decir, a través del objetivo). Estos microscopios se utilizan ampliamente en biología y son la base para diseños de microscopios más avanzados, como el microscopio confocal y el microscopio de fluorescencia de reflexión interna total (TIRF).
Microscopía de epifluorescencia

La mayoría de los microscopios de fluorescencia, especialmente los utilizados en las ciencias biológicas, tienen el diseño de epifluorescencia que se muestra en el diagrama. La luz de la longitud de onda de excitación ilumina la muestra a través de la lente del objetivo. La fluorescencia emitida por la muestra se enfoca al detector mediante el mismo objetivo que se utiliza para la excitación, el cual para mayor resolución necesitará lentes objetivo con mayor apertura numérica. Dado que la mayor parte de la luz de excitación se transmite a través de la muestra, sólo la luz excitadora reflejada llega al objetivo junto con la luz emitida y, por lo tanto, el método de epifluorescencia proporciona una alta relación señal-ruido. El divisor de haz dicroico actúa como un filtro de longitud de onda específica, transmitiendo luz fluorescente al ocular o detector, pero reflejando cualquier luz de excitación restante hacia la fuente.
Fuentes de luz
La microscopía de fluorescencia requiere una iluminación intensa, casi monocromática, que algunas fuentes de luz muy extendidas, como las lámparas halógenas, no pueden proporcionar. Se utilizan cuatro tipos principales de fuentes de luz, incluidas lámparas de arco de xenón o lámparas de vapor de mercurio con filtro de excitación, láseres, fuentes supercontinuas y LED de alta potencia. Los láseres se utilizan más ampliamente para técnicas de microscopía de fluorescencia más complejas, como la microscopía confocal y la microscopía de fluorescencia de reflexión interna total, mientras que las lámparas de xenón, las lámparas de mercurio y los LED con un filtro de excitación dicroico se utilizan comúnmente para los microscopios de epifluorescencia de campo amplio. Al colocar dos conjuntos de microlentes en la trayectoria de iluminación de un microscopio de epifluorescencia de campo amplio, se puede lograr una iluminación altamente uniforme con un coeficiente de variación del 1-2%.
Preparación de muestras
Pantallas superpuestas de cuatro canales fluorescentes
b) Cyan: [PLL-A546 fluorescencia] - contador genérico para visualizar superficies de células eucariotas
(c) Azul: [ fluorescencia del santuario] - manchas de ADN, identifica núcleos
d) Rojo: [autofluorescencia de clorofila] - resuelve cloroplastos

Para que una muestra sea adecuada para microscopía de fluorescencia, debe ser fluorescente. Existen varios métodos para crear una muestra fluorescente; las principales técnicas son el marcaje con tinciones fluorescentes o, en el caso de muestras biológicas, la expresión de una proteína fluorescente. Alternativamente, se puede utilizar la fluorescencia intrínseca de una muestra (es decir, autofluorescencia). En las ciencias biológicas, la microscopía de fluorescencia es una herramienta poderosa que permite la tinción específica y sensible de una muestra para detectar la distribución de proteínas u otras moléculas de interés. Como resultado, existe una amplia gama de técnicas para la tinción fluorescente de muestras biológicas.
Tintes biológicos fluorescentes
Se han diseñado muchos tintes fluorescentes para una variedad de moléculas biológicas. Algunas de ellas son moléculas pequeñas que son intrínsecamente fluorescentes y se unen a una molécula biológica de interés. Los principales ejemplos de estos son las tinciones de ácido nucleico como DAPI y Hoechst (excitadas por luz de longitud de onda ultravioleta) y DRAQ5 y DRAQ7 (excitadas de manera óptima por luz roja), que se unen al surco menor del ADN, etiquetando así los núcleos de las células. Otros son fármacos, toxinas o péptidos que se unen a estructuras celulares específicas y se han derivatizado con un indicador fluorescente. Un ejemplo importante de esta clase de tinción fluorescente es la faloidina, que se utiliza para teñir fibras de actina en células de mamíferos. Un nuevo péptido, conocido como péptido hibridador de colágeno, también puede conjugarse con fluoróforos y usarse para teñir fibras de colágeno desnaturalizadas. La tinción de las paredes celulares de las plantas se realiza mediante tintes o tintes que se unen a la celulosa o la pectina. La búsqueda de sondas fluorescentes con una alta especificidad que también permitan obtener imágenes en vivo de células vegetales está en curso.
Hay muchas moléculas fluorescentes llamadas fluoróforos o fluorocromos, como la fluoresceína, Alexa Fluors o DyLight 488, que pueden unirse químicamente a una molécula diferente que se une al objetivo de interés dentro de la muestra.
Inmunofluorescencia
La inmunofluorescencia es una técnica que utiliza la unión altamente específica de un anticuerpo a su antígeno para marcar proteínas específicas u otras moléculas dentro de la célula. Se trata una muestra con un anticuerpo primario específico para la molécula de interés. Un fluoróforo se puede conjugar directamente con el anticuerpo primario. Alternativamente, se puede utilizar un anticuerpo secundario, conjugado con un fluoróforo, que se una específicamente al primer anticuerpo. Por ejemplo, un anticuerpo primario generado en un ratón que reconoce la tubulina combinado con un anticuerpo secundario anti-ratón derivatizado con un fluoróforo podría usarse para marcar microtúbulos en una célula.
Proteínas fluorescentes
La comprensión moderna de la genética y las técnicas disponibles para modificar el ADN permiten a los científicos modificar genéticamente proteínas para que también lleven una proteína indicadora fluorescente. En muestras biológicas, esto permite a un científico hacer directamente fluorescente una proteína de interés. Luego se puede rastrear directamente la ubicación de las proteínas, incluso en células vivas.
Limitaciones
Los fluoróforos pierden su capacidad de emitir fluorescencia cuando se iluminan en un proceso llamado fotoblanqueo. El fotoblanqueo se produce cuando las moléculas fluorescentes acumulan daño químico de los electrones excitados durante la fluorescencia. El fotoblanqueo puede limitar gravemente el tiempo durante el cual se puede observar una muestra mediante microscopía de fluorescencia. Existen varias técnicas para reducir el fotoblanqueo, como el uso de fluoróforos más robustos, minimizando la iluminación o utilizando productos químicos fotoprotectores.
La microscopía de fluorescencia con proteínas indicadoras fluorescentes ha permitido el análisis de células vivas mediante microscopía de fluorescencia; sin embargo, las células son susceptibles a la fototoxicidad, particularmente con luz de longitud de onda corta. Además, las moléculas fluorescentes tienden a generar especies químicas reactivas cuando están bajo iluminación, lo que mejora el efecto fototóxico.
A diferencia de las técnicas de microscopía de luz transmitida y reflejada, la microscopía de fluorescencia solo permite la observación de estructuras específicas que han sido marcadas para fluorescencia. Por ejemplo, observar una muestra de tejido preparada con una tinción de ADN fluorescente mediante microscopía de fluorescencia sólo revela la organización del ADN dentro de las células y no revela nada más sobre la morfología celular.
Las técnicas computacionales que proponen estimar la señal fluorescente a partir de imágenes no fluorescentes (como el campo claro) pueden reducir estas preocupaciones. En general, estos enfoques implican entrenar una red neuronal convolucional profunda en células teñidas y luego estimar la fluorescencia en muestras no teñidas. Por lo tanto, al desacoplar las células bajo investigación de las células utilizadas para entrenar la red, las imágenes se pueden realizar más rápido y con una fototoxicidad reducida.
Técnicas de subdifracción
La naturaleza ondulatoria de la luz limita el tamaño del punto en el que se puede enfocar la luz debido al límite de difracción. Esta limitación fue descrita en el siglo XIX por Ernst Abbe y "limita la resolución de un microscopio óptico a aproximadamente la mitad de la longitud de onda de la luz utilizada". La microscopía de fluorescencia es fundamental para muchas técnicas que pretenden superar este límite mediante configuraciones ópticas especializadas.
En el siglo XX se han inventado varias mejoras en las técnicas de microscopía que han dado lugar a un aumento de la resolución y el contraste hasta cierto punto. Sin embargo, no superaron el límite de difracción. En 1978 se desarrollaron las primeras ideas teóricas para romper esta barrera mediante el uso de un microscopio 4Pi como microscopio de fluorescencia de barrido láser confocal donde la luz se enfoca idealmente desde todos los lados hacia un foco común que se utiliza para escanear el objeto por "puntos". -por punto' excitación combinada con excitación 'punto por punto' detección. Sin embargo, la primera demostración experimental del microscopio 4pi tuvo lugar en 1994. La microscopía 4Pi maximiza la cantidad de direcciones de enfoque disponibles mediante el uso de dos lentes objetivo opuestos o microscopía de excitación de dos fotones utilizando luz desplazada al rojo y excitación multifotónica.
La microscopía correlativa integrada combina un microscopio de fluorescencia con un microscopio electrónico. Esto permite visualizar la ultraestructura y la información contextual con el microscopio electrónico mientras se utilizan los datos del microscopio de fluorescencia como herramienta de etiquetado.
La primera técnica para lograr realmente una resolución de subdiffraction fue la microscopía STED, propuesta en 1994. Este método y todas las técnicas siguiendo el concepto RESOLFT dependen de una fuerte interacción no lineal entre moléculas de luz y fluorescente. Las moléculas son impulsadas fuertemente entre los estados moleculares distinguibles en cada ubicación específica, para que finalmente la luz pueda ser emitida a sólo una pequeña fracción del espacio, por lo tanto una resolución mayor.
Además, en la década de 1990 se desarrolló otro método de microscopía de súper resolución basado en la microscopía de campo amplio. Se logró una resolución de tamaño sustancialmente mejorada de nanoestructuras celulares teñidas con un marcador fluorescente mediante el desarrollo de la microscopía de localización SPDM y la iluminación láser estructurada (iluminación espacialmente modulada, SMI). La combinación del principio de SPDM con SMI dio como resultado el desarrollo del microscopio Vertico SMI. La detección de una sola molécula de colorantes fluorescentes con parpadeo normal, como la proteína fluorescente verde (GFP), se puede lograr utilizando un desarrollo posterior del SPDM, la llamada tecnología SPDMphymod, que permite detectar y contar dos tipos diferentes de moléculas fluorescentes a nivel molecular (esto Esta tecnología se conoce como microscopía de localización de dos colores o 2CLM).
Como alternativa, la llegada de la microscopía de localización fotoactivada podría lograr resultados similares basándose en el parpadeo o el cambio de moléculas individuales, donde la fracción de moléculas fluorescentes es muy pequeña en cada momento. Esta respuesta estocástica de las moléculas a la luz aplicada corresponde también a una interacción altamente no lineal, que conduce a una resolución de subdifracción.
Galería de micrografías de fluorescencia
 Una proyección z de una célula osteosarcoma, manchada con falloidina para visualizar filamentos de actina. La imagen fue tomada en un microscopio confocal, y la posterior desconvolución se hizo utilizando una función de diseminación de punto derivada experimentalmente.
Una proyección z de una célula osteosarcoma, manchada con falloidina para visualizar filamentos de actina. La imagen fue tomada en un microscopio confocal, y la posterior desconvolución se hizo utilizando una función de diseminación de punto derivada experimentalmente.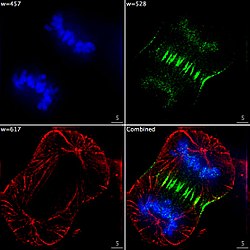 Imagen epifluorescente de los tres componentes en una célula de cáncer humano divisoria. El ADN está manchado de azul, una proteína llamada INCENP es verde, y los microtubulos son rojos. Cada fluoróforo se imagena por separado utilizando una combinación diferente de filtros de excitación y emisión, y las imágenes se capturan secuencialmente utilizando una cámara CCD digital, luego superpuesta para dar una imagen completa.
Imagen epifluorescente de los tres componentes en una célula de cáncer humano divisoria. El ADN está manchado de azul, una proteína llamada INCENP es verde, y los microtubulos son rojos. Cada fluoróforo se imagena por separado utilizando una combinación diferente de filtros de excitación y emisión, y las imágenes se capturan secuencialmente utilizando una cámara CCD digital, luego superpuesta para dar una imagen completa. Células endoteliales bajo el microscopio. Nuclei se manchan azul con DAPI, los microtubulos están marcados verde por un anticuerpo ligado a la FITC y los filamentos de actina se etiquetan rojo con phalloidin ligados a TRITC. Células endoteliales de arteria pulmonar bovina (BPAE)
Células endoteliales bajo el microscopio. Nuclei se manchan azul con DAPI, los microtubulos están marcados verde por un anticuerpo ligado a la FITC y los filamentos de actina se etiquetan rojo con phalloidin ligados a TRITC. Células endoteliales de arteria pulmonar bovina (BPAE)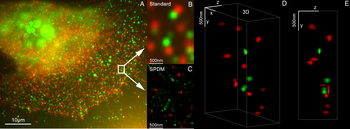 Microscopia de doble color 3D con Her2 y Her3 en células mamarias, tintes estándar: Alexa 488, Alexa 568. Microscopia LIMON
Microscopia de doble color 3D con Her2 y Her3 en células mamarias, tintes estándar: Alexa 488, Alexa 568. Microscopia LIMON núcleo de linfocitos humanos manchado con DAPI con cromosoma 13 (verde) y 21 (rojo) sondas centromere híbridas (hibridación in situ fluorescente)
núcleo de linfocitos humanos manchado con DAPI con cromosoma 13 (verde) y 21 (rojo) sondas centromere híbridas (hibridación in situ fluorescente) Membrana de células de levadura visualizada por algunas proteínas de membrana fusionadas con marcadores fluorescentes RFP y GFP. Imposición de luz de ambos marcadores resulta en color amarillo.
Membrana de células de levadura visualizada por algunas proteínas de membrana fusionadas con marcadores fluorescentes RFP y GFP. Imposición de luz de ambos marcadores resulta en color amarillo. Microscopia de Super-resolución: Detección de moléculas individuales de YFP en una célula de cáncer humano. Mediciones de distancia típicas en el rango de 15 nm medido con un microscopio Vertico-SMI/SPDMphymod
Microscopia de Super-resolución: Detección de moléculas individuales de YFP en una célula de cáncer humano. Mediciones de distancia típicas en el rango de 15 nm medido con un microscopio Vertico-SMI/SPDMphymod Microscopia de súper resolución: microscopia de colocalización (2CLM) con proteínas de fusión GFP y RFP (núcleo de una célula de cáncer de hueso) 120.000 moléculas localizadas en un área de amplio campo (470 μm2) medido con un microscopio Vertico-SMI/SPDMphymod
Microscopia de súper resolución: microscopia de colocalización (2CLM) con proteínas de fusión GFP y RFP (núcleo de una célula de cáncer de hueso) 120.000 moléculas localizadas en un área de amplio campo (470 μm2) medido con un microscopio Vertico-SMI/SPDMphymod Microscopia de fluorescencia de la Expresión de ADN en el Hombre Wild-Type y P239S Mutant Palladin.
Microscopia de fluorescencia de la Expresión de ADN en el Hombre Wild-Type y P239S Mutant Palladin. Imágenes de microscopía de fluorescencia de la patología del sol en una célula sanguínea mostrando las áreas afectadas en rojo.
Imágenes de microscopía de fluorescencia de la patología del sol en una célula sanguínea mostrando las áreas afectadas en rojo.










Contenido relacionado
Julio (unidad)
Ley de Fick
Pascal (unidad)