
Linfopoyesis
La linfopoyesis (lĭm'fō-poi-ē'sĭs) (o linfocitopoyesis) es la generación de linfocitos, uno de los cinco tipos de glóbulos blancos (GB). Se la conoce más formalmente como hematopoyesis linfoide.
La alteración de la linfopoyesis puede provocar una serie de trastornos linfoproliferativos, como linfomas y leucemias linfoides.
| Glosario de linfopoiesis |
|---|
| • antígeno cualquier molécula que pueda provocar una defensa inmune |
| • Células B linfocitos que finalmente producen anticuerpos |
| • médula ósea el centro de los huesos capaces de producir todos los glóbulos rojos y blancos en el adulto |
| • Corteza la porción externa de cualquier órgano |
| • citoplasma la porción de una célula entre el núcleo y la membrana |
| • diferenciación cambios permanentes en una célula que se desarrolla con el tiempo y con la división celular |
| • gránulos granos encontrados en muchos glóbulos blancos, compuestos de químicos defensivos |
| • hematopoyético que da lugar a cualquier tipo de célula sanguínea |
| • linaje un tipo de célula y sus descendientes por división y diferenciación |
| • linfocitos un 'lineaje' especial de WBC |
| • macrófagos descendientes de mieloide (algunos pueden ser linfoides) con habilidades de "comida", también cooperan con los linfocitos |
| • mieloide ancestros de WBC con gránulos y también de macrófagos |
| • T Cells linfocitos de "gestión" para inmunidad |
| • (WBC) White Blood Cell en contraste con la Célula de Sangre Roja mucho más común; responsable de la defensa |
| edición |
Terminología
Los linfocitos son células sanguíneas de linaje linfoide (en lugar de mieloide o eritroide).

Los linfocitos se encuentran en el torrente sanguíneo y tienen su origen en la médula ósea, aunque pertenecen principalmente al sistema linfático independiente, que interactúa con la circulación sanguínea.
En la actualidad, el término linfopoyesis se suele utilizar indistintamente con el de "linfocitopoyesis", es decir, la creación de linfocitos, pero algunas fuentes distinguen entre ambos términos y afirman que "linfopoyesis" también se refiere a la creación de tejido linfático, mientras que "linfocitopoyesis" se refiere únicamente a la creación de células en ese tejido. Actualmente, es poco frecuente que el término linfopoyesis se refiera a la creación de tejidos linfáticos.
Mielopoyesis se refiere a la "generación de células del linaje mieloide" y eritropoyesis se refiere a la "generación de células del linaje eritroide", por lo que se ha desarrollado un uso paralelo en el que linfopoyesis se refiere a la "generación de células del linaje linfoide".
Las dos clases de glóbulos blancos en ratones se originan a partir de células con fuertes propiedades de células madre: las mieloides, del progenitor mieloide común (CMP), y las linfoides, del progenitor linfoide común (CLP). Finalmente, se descubrió que estos progenitores no eran únicos y que las clases mieloide y linfoide no estaban separadas, sino que eran dos árboles genealógicos parcialmente entrelazados.
Función
Los linfocitos maduros son una parte fundamental del sistema inmunológico que, con excepción de las células B y T de memoria, tienen vidas cortas que se miden en días o semanas y deben generarse continuamente durante toda la vida mediante la división celular y la diferenciación a partir de células como los progenitores linfoides comunes (CLP) en ratones.
El conjunto de células CLP y progenitores similares son en sí mismos descendientes de la célula madre hematopoyética pluripotencial (pHSC), que es capaz de generar todos los tipos celulares del sistema completo de células sanguíneas. A pesar de su capacidad para generar el conjunto completo de linfocitos, la mayoría de los progenitores no son células madre verdaderas y deben renovarse continuamente mediante la diferenciación a partir de la célula madre pHSC.
Muchas células progenitoras también se denominan células de tránsito, a veces también llamadas células amplificadoras de tránsito, lo que significa que la célula de tránsito puede encontrar un nuevo sublinaje, pero el número de células resultantes es estrictamente limitado (aunque posiblemente muy grande, incluso billones, pero finito) y el linaje termina con células que mueren (por apoptosis) o permanecen como células que ya no pueden dividirse. Ejemplos de tales células son las UFC (unidades formadoras de colonias, denominadas así por su capacidad de formar colonias in vitro en medios artificiales) como las UFC-T.
El trasplante de una sola célula pHSC puede reconstituir un huésped irradiado de forma subletal (es decir, un ratón que ha sido irradiado de forma que se han destruido todos los leucocitos) con todos estos linajes de células, incluidos todos los tipos de linfocitos a través de las CLP.
La linfopoyesis continúa durante toda la vida, por lo que las células progenitoras y sus células madre progenitoras deben estar siempre presentes.
Sinopsis
En el caso de mamíferos como el ser humano (Homo sapiens), la linfopoyesis comienza con un aporte pasivo limitado por parte de la madre. Este incluye linfocitos e inmunoglobulina G que atraviesan la placenta y entran al feto para brindar cierta protección contra patógenos, así como leucocitos que provienen de la leche materna y entran a la circulación a través del tracto digestivo. A menudo no es eficaz para prevenir infecciones en el recién nacido.
Sin embargo, al principio de la gestación, el embrión en desarrollo ha iniciado su propia linfopoyesis a partir del hígado fetal. La linfopoyesis también surge del saco vitelino, a diferencia del adulto, donde todos los linfocitos se originan en la médula ósea.
Existen cuatro tipos principales de linfocitos, junto con muchos subtipos. Los científicos han identificado cientos o miles de tipos de células linfocitarias, todas ellas generadas por linfopoyesis normal o anormal, a excepción de ciertas cepas artificiales creadas en laboratorios mediante el desarrollo de cepas existentes. Aunque los linfocitos suelen considerarse maduros, como se observa en los análisis de sangre, ciertamente no son inertes. Los linfocitos pueden viajar por el cuerpo donde sea necesario. Cuando surgen tales necesidades, pueden producirse nuevas rondas de linfopoyesis descendentes, como la multiplicación y diferenciación celular, acompañadas de una intensa actividad mitótica y metabólica.
No se trata de un tema sencillo. En su texto de 1976 Inmunología, envejecimiento y cáncer, el inmunólogo y premio Nobel Sir Frank Macfarlane Burnet especuló que algún día se descubriría que el sistema inmunológico es tan complejo como el sistema nervioso. Como la producción de linfocitos está tan cerca del papel central de la respuesta inmunológica, es prudente abordar su estudio con cierta humildad ante la tarea. Sin embargo, existen principios generales que ayudan a comprenderlo.
Proceso
La linfopoyesis puede considerarse en un sentido matemático como un proceso recursivo de división celular y también como un proceso de diferenciación, medido por cambios en las propiedades de las células.
- Dado que los linfocitos surgen de tipos específicos de células madre limitadas – que podemos llamar P (para Progenitor) células – tales células pueden dividirse de varias maneras. Estos son principios generales de células madre limitadas.
Si consideramos a la P como la célula «madre», pero no como una célula madre verdadera, puede dividirse en dos nuevas células, que son idénticas, pero difieren en algún grado de la madre. O la célula madre P puede dividirse de manera desigual en dos nuevas células hijas, que difieren entre sí y también de la madre.
Cualquier célula hija normalmente tendrá nuevas capacidades especializadas y si es capaz de dividirse formará un nuevo sublinaje. La diferencia entre una célula hija y la madre puede ser grande, pero también puede ser mucho menor, incluso sutil. Lo que la célula madre P no hace es dividirse en dos nuevas células madre P o en una madre y una hija; esto es una cuestión de observación, ya que se sabe que estas células progenitoras limitadas no se auto-renuevan.
- Hay una especie de excepción cuando las células hijas en algún nivel del linaje pueden dividir varias veces para formar células más aparentemente idénticas, pero entonces inevitablemente se producirá una nueva diferenciación y división, hasta que se llegue a una etapa final en la que no puede ocurrir ninguna división más, y el linaje tipo célula es finalmente maduro. Un ejemplo de madurez es una célula plasmática, del linaje celular B, que produce anticuerpo copioso, pero no puede dividir y finalmente muere después de unos días o semanas.
- El progenitor CLP del ratón o el progenitor MLP del ser humano se diferencia en linfocitos convirtiéndose en un linoblasto (inmunología médica, p.10). Luego divide varias veces más para convertirse en un prolymphocyte que tiene marcadores específicos de la superficie celular únicos a una célula T (1) o (2) B. El progenitor también puede diferenciarse en (3) células asesinas naturales (NK) y (4) células dendritas (DC).
- Las células T, las células B y las células NK (y todas las demás células linfoides Innate) son únicas para la familia linfocitaria, pero las células dendritas no lo son. DC que son de apariencia idéntica pero tienen diferentes marcadores se diseminan a través del cuerpo, y provienen de linajes linfoides y mieloides. Sin embargo, estas células pueden tener tareas algo diferentes y pueden tomar alojamiento preferentemente en diferentes lugares. (Revise in light of new research) Esta es ahora una pregunta abierta; también, los diferentes linajes dendriáticos pueden tener diferentes ‘taks’ o funciones y permanecer en diferentes ‘localizaciones. ’
Los linfocitos T y B son indistinguibles bajo el microscopio. Las células B y T inactivas son tan anodinas, con pocos orgánulos citoplasmáticos y cromatina mayoritariamente inactiva, que hasta la década de 1960 los libros de texto podían describir estas células, que ahora son el foco central de la inmunología, como si no tuvieran ninguna función conocida.
Sin embargo, los linfocitos T y B son linajes celulares muy distintos y “crecen” en diferentes lugares del cuerpo. Desempeñan funciones muy diferentes (aunque cooperativas) en el organismo. Nunca se ha encontrado evidencia de que las células T y B puedan interconvertirse. Las células T y B son bioquímicamente distintas y esto se refleja en los diferentes marcadores y receptores que poseen en sus superficies celulares. Esto parece ser cierto en todos los vertebrados, aunque existen muchas diferencias en los detalles entre las especies.
- Independientemente de si la CLP (museo) o MLP o un pequeño conjunto de células progenitoras estrechamente relacionadas tienen crédito para generar la profusión de linfocitos, los mismos progenitores linfoides todavía pueden generar algunas células claramente mieloides.
Lymphopoiesis para Células
Las células T se forman en la médula ósea y luego migran a la corteza del timo para madurar en un entorno libre de antígenos durante aproximadamente una semana, donde apenas un 2-4% de las células T logran hacerlo. El 96-98% restante de las células T mueren por apoptosis y son fagocitadas por los macrófagos en el timo. Muchos timocitos (células T) mueren durante el proceso de maduración porque se realiza un examen intensivo para asegurarse de que cada timocito pueda reconocer el complejo péptido-MHC propio y para demostrar su propia tolerancia. Después de experimentar la apoptosis, el timocito muere y se recicla rápidamente.
Al madurar, existen varias formas de timocitos, entre ellas:
- T-helper (necesitado para la activación de otras células como células B y macrófagos),
- T-citotóxico (que mata células viralmente infectadas),
- T-memoria (células T que recuerdan los antígenos previamente encontrados) y
- Células T-suppressor (que moderan la respuesta inmune de otros leucocitos). También se llama células reguladoras T (Treg)
Cuando las células T se activan, experimentan una serie de nuevos desarrollos. Un linfocito T pequeño en reposo sufre rápidamente una transformación blastogénica en un linfocito grande (13-15 μm). Este linfocito grande (conocido en este contexto como linfoblasto) se divide luego varias veces para producir una población ampliada de linfocitos medianos (9-12 μm) y pequeños (5-8 μm) con la misma especificidad antigénica. Los linfocitos T finalmente activados y diferenciados son nuevamente morfológicamente indistinguibles de un linfocito pequeño en reposo. Por lo tanto, los siguientes estados de desarrollo pueden observarse en secuencia en los análisis de sangre:
- Prolibfocito
- Linfocto grande
- Linfocto pequeño
Mapa básico de la linfopoyesis de células T
Este mapa básico de la formación de células T en secuencia es simplificado y se parece a las descripciones de los libros de texto, y puede no reflejar las últimas investigaciones. (Inmunología médica, pág. 119)
En el timo
- MLP
- ETP
- DN1
- (B; Mφ)
- DN2
- (DC; NK)
- DN3
- (γδ)
- DN4
- DP
- (TNK; CD4; CD8; Treg)
En la periferia
- (Th1; Th2)
Desarrollo de células T
A diferencia de otros linajes linfoides, el desarrollo de las células T se produce casi exclusivamente en el timo. La linfopoyesis de células T no se produce de forma automática, sino que requiere señales generadas por las células del estroma del timo. Se han definido varias etapas en las que se requieren reguladores y factores de crecimiento específicos para que se produzca el desarrollo de las células T. Más adelante, en el desarrollo de las células T y su maduración, estos mismos factores reguladores se utilizan nuevamente para influir en la especialización de las células T.
Las células T son únicas entre las poblaciones de linfocitos por su capacidad de especializarse aún más como células maduras y volverse aún más maduras. Las células T se presentan en muchos tipos, por ejemplo: las células T TcRαβ convencionales; las llamadas células T TcRγδ no convencionales; las células NKT; y las células T reguladoras (Treg). Los detalles sobre el desarrollo y el ciclo de vida de las células T no convencionales están menos bien descritos en comparación con las células T convencionales.
Etapas de la maduración de las células T
Primera etapa: Migración del timo
Los progenitores linfoides multipotentes (MLP) ingresan a la vía de las células T a medida que migran al timo. Las células más primitivas del timo son los progenitores timocitos tempranos (ETP), que conservan todo el potencial linfoide y mieloide pero existen solo de manera transitoria, diferenciándose rápidamente en linajes T y NK. (Inmunología médica, pág. 118)
Etapa dos: Expansión proliferativa y compromiso del linaje T
El compromiso final con el linaje de las células T se produce dentro del microambiente del timo, las estructuras microscópicas del timo donde se nutren las células T. Las células T más primitivas conservan la capacidad multipotencial y pueden diferenciarse en células de los linajes mieloide o linfoide (células B, células dendríticas, células T o células NK).
Las células T doble negativas más diferenciadas (células DN2) tienen un potencial más limitado, pero aún no están completamente restringidas al linaje de células T (aún pueden convertirse en células dendríticas, células T o células NK). Más adelante, están completamente comprometidas con el linaje de células T: cuando los timocitos que expresan receptores Notch1 se unen a las células del estroma del timo que expresan ligandos Notch1, los timocitos finalmente se comprometen con el linaje de células T. Ver la imagen de la galería "Doble negativas"
Con el compromiso con el linaje de células T, comienza un proceso muy complejo conocido como reordenamiento del gen TCR. Esto crea una enorme diversidad de células T portadoras de receptores de antígenos. Después, algunas células T abandonan el timo para migrar a la piel y las mucosas.
Tercera etapa: β-selección
Etapa cuatro: Selección de receptores de células T
Sólo entre el 2% y el 3% de los timocitos diferenciadores, aquellos que expresan TcR capaces de interactuar con moléculas MHC, pero tolerantes a péptidos propios, sobreviven al proceso de selección de la Etapa Cuatro.
Quinta etapa: Diferenciación continua en la periferia
Anteriormente se creía que el timo humano permanecía activo como sitio de diferenciación de células T sólo hasta la edad adulta temprana y que más tarde en la vida adulta el timo se atrofia, quizás incluso desapareciendo. Informes recientes indican que el timo humano está activo durante toda la vida adulta. Por lo tanto, varios factores pueden contribuir al suministro de células T en la vida adulta: generación en el timo, diferenciación extratímica y el hecho de que las células T de memoria son longevas y sobreviven durante décadas.
Tipos de células T
- No convencional Células
El timo también da origen a las llamadas "células T no convencionales", como las células T γδ, las células T asesinas naturales (NKT) y las células T reguladoras (Treg).
- γδ células T
Las células γδT representan solo entre el 1% y el 5% de las células T circulantes, pero son abundantes en el sistema inmunitario de las mucosas y en la piel, donde representan la población dominante de células T. Estas "células T no restringidas por el MHC" participan en respuestas inmunitarias primarias específicas, vigilancia tumoral, regulación inmunitaria y cicatrización de heridas.
Se han descrito varias diferencias entre el desarrollo de las células T αβ y γδ. Estas células emigran del timo en "olas" de poblaciones clonales, que se alojan en tejidos discretos. Por ejemplo, un tipo se encuentra en la sangre periférica mientras que otro predomina en el tracto intestinal.
- Asesino natural Células
Las células NKT humanas son una población única y se cree que desempeñan un papel importante en la inmunidad tumoral y la inmunorregulación.
- T Células reguladoras
Las células Treg se consideran células T reguladoras naturales. Las Treg constituyen aproximadamente el 5 % de las células T CD4+ circulantes. Se cree que estas células poseen una propiedad autoinmunitaria importante al regular las células T "autorreactivas" en la periferia. (Inmunología médica, pág. 117-122)
Linfopoiesis para células B
Las células B se forman y maduran en la médula ósea (y el bazo).
Es una buena ayuda mnemotécnica decir que las células B se forman en la médula ósea, pero es una mera coincidencia, ya que las células B se estudiaron por primera vez en la bolsa de Fabricio del pollo y es de esta bolsa de donde las células B obtienen su nombre.
Estas células B abandonan la médula ósea y migran a través del torrente sanguíneo y la linfa hacia los tejidos linfoides periféricos, como el bazo, los ganglios linfáticos, las amígdalas y los tejidos mucosos. Una vez en un órgano linfoide secundario, la célula B puede ser introducida en un antígeno que es capaz de reconocer.
A través de este reconocimiento de antígenos y otras interacciones celulares, la célula B se activa y luego se divide y se diferencia para convertirse en una célula plasmática. La célula plasmática, un producto final de la célula B, es una célula secretora de anticuerpos muy activa que ayuda a proteger el cuerpo al atacar y unirse al antígeno.
Incluso después de muchas décadas de investigación, sigue habiendo cierta controversia sobre dónde maduran y "completan su educación" las células B, aunque existe la posibilidad de que el sitio también pueda estar parcialmente formado por tejidos linfoides periintestinales.
La linfopoyesis B ocurre exclusivamente en la médula ósea y los linfocitos B se producen allí continuamente durante toda la vida en un "microambiente" compuesto de células del estroma, matriz extracelular, citocinas y factores de crecimiento, que son fundamentales para la proliferación, diferenciación y supervivencia de los linfocitos tempranos y los precursores del linaje B.
La proporción relativa de células B precursoras en la médula ósea permanece bastante constante a lo largo de la vida de un organismo. Existen estadios como las células Pre-B-I (del 5% al 10% del total), las células Pre-B-II (del 60% al 70%) y el 20% al 25% restante son células B inmaduras. La mayoría de los libros de texto dicen que las células B maduran en la médula ósea, pero, por lo general, las células B inmaduras migran al bazo para recibir algún tipo de "educación superior", donde pasan por estadios de transición antes de la maduración final. (Inmunología médica, pág. 136)
Los linfocitos B se identifican por la presencia de inmunoglobulina G soluble (IgG). Esta es la inmunoglobulina protectora más común en el cuerpo adulto. Después de la estimulación antigénica, las células B se diferencian en células plasmáticas que secretan grandes cantidades de IgG soluble. Esta es la etapa final de la linfopoyesis B, pero es la decisiva porque las células plasmáticas deben emitir anticuerpos cerca de una fuente de infección o diseminarlos en la sangre para combatir una infección a distancia o en una parte inaccesible del cuerpo.
Mapa básico de la linfopoyesis de células B

Un mapa generalmente considerado válido de la linfopoyesis de las células B es el siguiente en secuencia, en dos partes, la primera en la médula ósea y la segunda en el bazo: El proceso de desarrollo en la médula ósea ocurre en los centros germinales.
En la médula ósea
- Pro-B
- Pre-B-I
- Pre-B-II grande
- Pre-B-II pequeño
- Imm(ature)
En el bazo
- T1
- T2/T3
- (Zona marginal); B-1; B-2)
- B-2 también se diferencian en:
- (Germinal Center (GC); Memory; Plasma)
Linfopoiesis para células NK
Las células NK, que carecen de receptores específicos de antígenos, se desarrollan en la médula ósea. Después de la maduración y liberación de la médula, circulan en la sangre durante toda su vida en busca de oportunidades. La oportunidad que buscan es encontrar, reconocer y luego matar células anormales como células cancerosas o células infectadas por virus. Es bien sabido que los linfocitos nunca tienen gránulos o al menos no gránulos que sean fácilmente visibles incluso al teñirlos. Sin embargo, las células NK son la excepción. Tienen numerosos gránulos que les proporcionan la capacidad de matar células y estos gránulos son la razón por la que las células NK tienen un nombre alternativo: linfocitos granulares grandes (LGL).
Las células NK son los únicos linfocitos considerados parte del sistema inmunitario innato (en contraste con el sistema inmunitario adaptativo). Sin embargo, están mucho más relacionadas con las células T (parte del sistema inmunitario adaptativo) que con otras células del sistema inmunitario innato. Las células NK no solo comparten muchos marcadores de superficie, funciones y actividades en común con las células T, sino que también surgen de un progenitor T/NK común. También se cree que el precursor T/NK es la fuente de una subpoblación de células dendríticas linfoides. (Inmunología médica, pág. 121)
Las células NK tienen una definición de linfocitos CD3, CD16+, CD56t (consulte la sección de códigos de barras de este artículo). Los progenitores de NK se pueden encontrar principalmente en el timo (ratón), pero el timo no es absolutamente necesario para el desarrollo de las células NK. Es probable que las células NK puedan desarrollarse en una variedad de órganos, pero no se conoce el sitio principal de desarrollo de las células NK.
En los seres humanos, la mayoría (85-90%) de las células NK tienen una alta capacidad citolítica (la capacidad de lisar células). Un subconjunto más pequeño (10-15%) llamado NK "CD56 brillante" es el principal responsable de la producción de citocinas y tiene una mayor supervivencia. Al viajar a los ganglios linfáticos, las células NK "CD56 brillante" se diferencian nuevamente en células NK maduras que expresan receptores similares a inmunoglobulina de células asesinas (KIR), receptores de citotoxicidad natural (NCR) y moléculas de adhesión críticas. (Inmunología médica, pág. 122)
Linfopoiesis para células dendritas
El proceso por el cual las células CLP pueden diferenciarse para generar células dendríticas de linaje linfoide aún no está bien definido.
Las células dendríticas (DC) son células presentadoras de antígenos altamente especializadas y eficientes. Las células de apariencia idéntica provienen tanto de un linaje mieloide (denominadas células dendríticas mieloides) como de un linaje linfoide (denominadas células dendríticas plasmocitoides).
El desarrollo y la regulación de las células dendríticas no están bien caracterizados. Si bien se han identificado precursores de células dendríticas en el hígado, el timo y la médula ósea de fetos humanos, se cree que durante la vida adulta las células dendríticas se producen únicamente en la médula ósea y se liberan en la sangre para que vaguen y se asienten. En general, se distribuye una gran cantidad de células dendríticas de diversos tipos por todo el cuerpo, especialmente en epitelios como la piel, para controlar a los invasores y mordisquear sus antígenos. (Inmunología médica, pág. 122)
Comparación de asesinos de linfocitosis
Los linfocitos tienen una serie de propiedades alarmantes, como la capacidad de deambular por el cuerpo y alojarse prácticamente en cualquier lugar, y mientras están en el camino emiten órdenes en forma de citocinas, quimiocinas y linfocinas, órdenes que afectan a muchos tipos de células del cuerpo y que también pueden inducir recursivamente más linfopoyesis. Un patrón de comportamiento fuerte que cautiva a los investigadores y al público por igual es la capacidad de los linfocitos de actuar como policía, juez y verdugo para matar a otras células o exigirles que se suiciden, una orden que generalmente se obedece. No parece haber otra opción de sentencia disponible.
Las células asesinas se distinguen de las células, como los macrófagos, que se comen a otras células o mastican desechos mediante un método llamado fagocitosis. Las células asesinas no utilizan la fagocitosis, sino que simplemente matan y dejan que otras células se encarguen de la limpieza.
Se sabe que los agentes asesinos atacan a las células infectadas por virus y a las células que se han vuelto cancerosas. Debido a estas capacidades, se han realizado muchas investigaciones para transformar estas cualidades en terapias médicas, pero el progreso ha sido lento.
Aquí está el desfile de asesinos y cómo trabajan:
- citotóxico Células
(también llamados Tc o linfocitos T citolíticos o tóxicos específicos de antígeno (CTL)). Los Tc matan por apoptosis y rocian su objetivo con perforina o granzimas o utilizan la interacción Fas-Fasl para ordenar la eliminación del objetivo. Esto mata las células que están infectadas y muestran antígeno.
- Células NK (también llamadas LGL ( linfocitos granulares grandes))
Matan con exactamente los mismos métodos que las células Tc pero no interactúan con ningún antígeno. Seleccionan sus objetivos en función de las moléculas típicas que presentan las células sometidas a estrés por una infección viral. Las células NK se encuentran principalmente en la circulación (5-15% de los linfocitos circulantes), pero también se distribuyen en los tejidos de todo el mundo.
- Las células LAK (Asesino activado por linfoquina) son un subconjunto laboratorio/clínico de células NK promovido por IL-2 para atacar células tumorales.
- Las células NKT ven asesino natural T cell main article
Células T asesinas naturales. Las células T NK humanas son una población única (que expresan marcadores de células NK como CD56 y KIR). Se cree que las células NKT desempeñan un papel importante en la inmunidad tumoral y la inmunorregulación (Medical Immunology, p. 135), pero se sabe poco al respecto. Evidencias recientes sugieren un papel en conjunto con las células estrelladas hepáticas, ya que son células presentadoras de antígenos residentes en el hígado que presentan antígenos lipídicos y estimulan la proliferación de las células NKT.
- Células T como asesino natural
Un grupo heterogéneo con propiedades mal definidas.
Sin embargo, en resumen, no existe ninguna célula o conjunto de células conocidas que sea capaz de matar las células cancerosas en general.
Linfopoiesis de etiqueta
Dado que todos los glóbulos blancos son microscópicos, incoloros y, a menudo, aparentemente idénticos en apariencia, se los identifica individualmente por sus marcadores químicos naturales, muchos de los cuales han sido analizados y nombrados. Cuando dos células tienen los mismos marcadores, se supone razonablemente que las células son idénticas en ese momento. Un conjunto de marcadores se describe coloquialmente como el código de barras de esa célula o de esa línea celular.
- He aquí un ejemplo de cómo puede llegar a ser un código de barras, para la célula hematopoyética totalmente importante (HSC) como ejemplo.
Las HSC se describen técnicamente como: carentes de la tirosina quinasa 3 similar a FMS (Flt3) y de los marcadores específicos de linajes linfoides discretos (Lin), pero que expresan altos niveles de Sca1 y c-kit; las HSC también expresan CD44, niveles bajos de Thy1.1 (CD90), pero no IL-7Ra ni CD27.
Esto se denomina fenotipo (superficial) de una HSC. Puede expresarse como un conjunto (Lin2, Sca1high, c-kit high, CD44+, Thy1.1low, CD27 2 e IL-7Ra2). Este conjunto es un «código de barras» para la HSC, similar a la etiqueta de código de barras que se coloca en la bolsa de plástico de las alas de pollo para pagar en el supermercado. Los científicos utilizan estos códigos de barras para verificar, categorizar y acumular células para muchos propósitos, a menudo utilizando métodos de laboratorio como la citometría de flujo celular. Estos códigos de barras definen parcialmente el significado moderno de fenotipo para los leucocitos.
La progresión de la diferenciación de las células madre hematopoyéticas y el compromiso de linaje se indican mediante cambios en este fenotipo. Es decir, a medida que la célula cambia, los marcadores también cambiarán y el código de barras cambiará.
- Códigos de barras típicos para algunos tipos de celda que aparecen en este artículo.
| Tipo de célula | Código de barras |
|---|---|
| ETP | C-Kit+, CD44+, CD25- |
| DN1 | CD44+, CD25- |
| DN2 | CD44+, CD25+ |
- Nota explicando los detalles del parámetro de código de barras: Flt3 es un receptor de cytokine tirosina kinase considerado importante en el desarrollo linfoides temprano. Además, Flt3 desempeña un papel importante en el mantenimiento de los progenitores linfoides B. CD27 juega un papel en la proliferación linfoides, diferenciación y apoptosis. La adquisición de CD27 y Flt3 por el HSC coincide con la pérdida de potencial de repoblación a largo plazo. En esta etapa las células conservan el potencial linfoides y mieloide y se denominan progenitores multipotentes. (Medical Immunology, p. 114)
Desarrollo del conocimiento con respecto a la linfapoiesis
Continuamente surgen nuevas preguntas en la inmunología, como si existiera una célula madre para ellas. Por ejemplo, se pensaba que el proceso de linfopoyesis era una secuencia unidireccional, ordenada y directa. Pero no está claro si los linfocitos en etapa terminal provienen de progenitores que son poblaciones homogéneas o poblaciones superpuestas. Tampoco está claro si los linajes de linfocitos se desarrollan a través de un continuo de diferenciación con una pérdida progresiva de opciones de linaje o si los eventos abruptos resultan en la adquisición de ciertas propiedades.
Los cambios en el citoplasma, la morfología del núcleo celular, los gránulos, la bioquímica interna celular, las moléculas de señalización y los marcadores de la superficie celular son difíciles de correlacionar con etapas definidas de la linfopoyesis. Las diferencias morfológicas no sólo corresponden a pasos de la mitosis (división de células somáticas), sino que son el resultado de continuos "procesos de maduración" del núcleo celular, así como del citoplasma, por lo que no se debe ser demasiado rígido en cuanto a las distinciones morfológicas entre ciertas etapas celulares.
- Modelos y actualizaciones sobre el árbol de la familia de la linfocitosis
Hasta hace poco, el modelo del CMP que genera todas las células mieloides y del CLP que genera todas las células linfoides se consideraba necesario y suficiente para explicar los hechos conocidos observados en la generación de glóbulos blancos, y todavía se encuentra en la mayoría de los libros de texto básicos. Sin embargo, a partir de alrededor de 2000 y ganando impulso después de 2005, tanto en estudios realizados en humanos como en ratones, se observaron nuevas complejidades que se publicaron en artículos. Estos estudios son importantes ahora principalmente para los investigadores de inmunología, pero es probable que con el tiempo conduzcan a cambios en los tratamientos médicos.
Los cambios fueron provocados por observaciones de que la linfopoyesis no siempre se dividía en dos linajes a nivel de los CLP. Peor aún, algunos macrófagos (considerados durante mucho tiempo un linaje mieloide) podían ser generados por progenitores del linaje linfoide. En esencia, el enfoque se ha desplazado del CLP a los MLP (progenitores específicos de linfoides), que son claramente progenitores linfoides pero conservan cierto potencial mieloide, en particular la capacidad, tanto en humanos como en ratones, de producir macrófagos (uno de los defensores de células inmunitarias más versátiles) y también muchas células dendríticas, los mejores "perros guardianes" de los invasores de antígenos.
Sin embargo, cualesquiera que sean los detalles, el proceso de linfopoyesis siempre parece generar incesantemente una progenie con atributos y capacidades especiales –"superpoderes" por así decirlo– pero con un potencial de desarrollo linfoide progresivamente más restringido.
Etapas del desarrollo
El viejo modelo: linfoides vs mieloide
Este modelo de linfopoyesis tenía la virtud de una relativa simplicidad, de concordancia con la nomenclatura y la terminología, y todavía es esencialmente válido para el ratón de laboratorio.
- pHSC pluripotente, auto-renovación, células madre hematopoyéticas que dan lugar a
- MPP progenitores multipotentes, que dan lugar a
- ELP (o PRO) Prolymphocytes, progenitores linfoides tempranos, y finalmente a los
- CLP progenitor linfoides común, un tipo de célula totalmente comprometido con el linaje linfoides.
Las células pHSc, MPP y ELP no están completamente comprometidas con el linaje linfoide porque si una de ellas se traslada a una ubicación diferente puede diferenciarse en una progenie no linfoide. Sin embargo, las CLP están comprometidas con el linaje linfoide. La CLP es la célula de tránsito responsable de estas etapas de desarrollo (generalmente paralelas), que se indican a continuación:
- Células NK
- Células dendritas ( linaje linfático; DC2)
- Progenitor Células B
- Células Pro-B Próxima Pro (o pre-pre)-B celdas = Células Pro tardías (o previas)-B
- Grandes células Pre-B = Pequeño pre- Células B
- Células Immature B
- Células B = coeficiente (células B1; células B2)
- Células de plasma
- Células Pro-T
- T-cells
Investigación sobre nuevos modelos (no ratones)
En 2008 se descubrió que "la mayoría de las células progenitoras tímicas tempranas [ETP] no se comprometen a convertirse en células T cuando llegan a la glándula del timo. Las células ETP conservan la capacidad de convertirse en células T o células mieloides".
Véase también:
Vista gráfica del modelo antiguo vs modelo linfoides mixto y mieloide
 A un lado. Comparando los modelos nuevos y antiguos de linaje.
A un lado. Comparando los modelos nuevos y antiguos de linaje.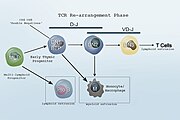 Linfoide linaje revisado y diagrama de flujo mieloide.
Linfoide linaje revisado y diagrama de flujo mieloide.
_Grayscale.jpg)

Textos de referencia general de inmunología
Los textos en negrita son los más citados en este artículo.
- Cell Communication in Nervous and Immune System; Gundelfinger, Seidenbecher, Schraven; Springer Berlin Heidelberg New York; 2006; ISBN 3-540-36828-0
- Color Atlas de Hematología; Theml et al.; Thieme; 2004; ISBN 1-58890-193-9
- Dinámica del cáncer; Steven A. Frank; Princeton University Press, Princeton, New Jersey; 2007; ISBN 978-0-691-13366-9, Creative Commons Public License
- Inmunología fundamental, 5a edición; William E. Paul (editor); Lippincott Williams & Wilkins Editores; 2003; ISBN 0-7817-3514-9
- Inmuno-Biología: El Sistema Inmunitario de Salud y Ciencia, 6a edición; Janeway, Travers; 2005; Garland Science Publishing, Nueva York; ISBN 0-8153-4101-6
- Introductor de inmunología Textbook (ebook;revised 2nd edition); Nandini Shetty; New Age International (P) Limited, Publishers, India; 2005; ISBN 81-224-2335-3
- Notas instantáneas en inmunología, 2a edición; Lydyard, Whelan, Fanger; Taylor y Francis Group; 2004; China Version ISBN 978-7-03-025225-8; 46RMB Wangfujing Bookstore
- Inmunología médica—6a edición; G. Virella, Editor; Informa Healthcare USA, Inc; 2007; ISBN 978-0-8493-9696-0
- Biología de células madre; Marshak, Gardner, Gottlieb; Cold Spring Harbor Laboratory Press; 2001; ISBN 0-87969-575-7/01
- Textbook of Human Development and Histology; Zhong Cuiping et al.; Shanghai Scientific and Technical Publishers; 2006; ISBN 7-5323-8230-3
- Textbook of Medical Immunology (Inmunología, 7a edición); LIM Pak Leong; Elsevier (Singapur) Pte Ltd.; 2006; ISBN 0-323-03399-7
Referencias
- ^ a b c d e Birbrair, Alexander; Frenette, Paul S. (2016-03-01). "Nicho heterogeneidad en la médula ósea". Annals of the New York Academy of Sciences. 1370 (1): 82–96. código:2016NYASA1370...82B. doi:10.1111/nyas.13016. ISSN 1749-6632. PMC 4938003. PMID 27015419.
- ^ Biología de células madre, página 307
- ^ Dinámica del cáncer, página 251
- ^ "CFU-T". Archivado desde el original el 2023-05-26. Retrieved 2010-08-24.
- ^ Reber AJ, Donovan DC, Gabbard J, Galland K, Aceves-Avila M, Holbert KA, Marshall L, Hurley DJ (2008). "La transferencia de leucocitos colostrales maternas promueve el desarrollo del sistema inmunológico neonatal Parte II. Efectos sobre linfocitos neonatales". Vet Immunol Immunopathol. 123 (3–4): 305–13. doi:10.1016/j.vetimm.2008.02.009. PMID 18394715.
- ^ Pryhuber, Gloria S. (2015). "Infecciones postnatales e inmunología que afecten la enfermedad crónica pulmonar de la prematuridad". Clínicas en Perinatología. 42 (4): 697–718. doi:10.1016/j.clp.2015.08.002. ISSN 0095-5108. PMC 4660246. PMID 26593074.
- ^ Progeniores de células T adultos retienen potencial mieloide; Naturaleza; 2008
- ^ Textbook of Human Development and Histology, p.176
- ^ Inmuno-Biología, Sistema Inmunitario de Salud y Ciencia.
- ^ Inmunología fundamental Quinta edición
- ^ Inmuno-Biología, Sistema Inmunitario de Salud y Ciencia. Garland Science Publishing
- ^ Ciofani, Maria; Zúñiga-Pflücker, Juan Carlos (2007-01-01-01). "El timo como un sitio inductivo para la linfopoiesis T". Annual Review of Cell and Developmental Biology. 23: 463–493. doi:10.1146/annurev.cellbio.23.090506.123547. ISSN 1081-0706. PMID 17506693. Archivado desde el original el 2021-09-03. Retrieved 2019-07-03.
- ^ Textbook of Medical Immunology, página 5
- ^ Inmunología médica, página 23
- ^ "Gamma Delta (γδ) T Cells durable British Society for Immunology". www.immunology.org. Archivado desde el original el 20 de diciembre de 2012. Retrieved 2019-06-14.
- ^ "Inmunidad tumoral y inmunoterapia del cáncer". Archivado desde el original el 2010-08-27. Retrieved 2010-08-25.
- ^ Inmunología médica, página 22
- ^ Inmunología médica, pág. 123
- ^ Inmunología fundamental; Paul; Ch. 15 "DISTRIBUTION OF DENDRITIC CELLS IN VIVO: A MULTIMEMBER FAMILY"
- ^ Inmunología; Lydyard et al; p. 22, 132-137
- ^ Inmunología; Lydyard et al; p. 15, 18-20,41
- ^ Inmunología; Lydyard et al; págs. 20, 259 a 260
- ^ Inmunología médicaLitwin, pág. 122
- ^ Color Atlas de Hematología 2004
- ^ "Los Progeniores Trómicos más antiguos para las células T Posee potencial de linaje mieloide"; Bell, Bhandoola; Vol 452, 10 de abril de 2008, doi:10.1038/nature06840
- ^ Inmunología médica, Litwin, p.115
- ^ Textbook of Medical Immunology, página 31
- ^ Búsquedas May Shed Light on T-cell Leucemias e Inmunodeficiencias. Archivado 2010-02-13 en el Wayback Machine Bhandoola. 9 de abril de 2008;
- ^ Líneas de Sangre Redrawn; Thomas Graf; Naturaleza Vol 452 10 April 2008 p.702-703
- ^ The Common Myelolymphoid Progenitor: A Key Intermediate Stage in Hemopoiesis Generating T and B Cells; Min Lu, Hiroshi Kawamoto, Yoshihiro Katsube, Tomokatsu Ikawa y Yoshimoto Katsura; J. Immunol. 2002; 169;3519-3525
- ^ Identificación de Flt3 + Lympho-Myeloid Células madre que carecen de Erythro-Megakaryocytic Potential: A Revised Road Map for Adult Blood Lineage Commitment; Lund Strategic Research Center for Stem Cell Biology; Cell; Vol. 121, 295–306, 22 de abril de 2005
- ^ Los progenitores adultos de células T conservan potencial mieloide; Haruka Wada, Kyoko Masuda, Rumi Satoh, Kiyokazu Kakugawa, Tomokatsu Ikawa, Yoshimoto Katsura & Hiroshi Kawamoto; Naturaleza Vol 452 10 abril 2008
- ^ Los primeros progenitores tímicos de las células T poseen potencial de linaje mieloide; J. Jeremiah Bell, Avinash Bhandoola; Naturaleza; Vol 452, 10 de abril de 2008, págs. 764 a 768
- ^ El mapa revisado de la jerarquía de los progenitores humanos muestra el origen de los macrófagos y las células dendritas en el desarrollo linfoides temprano; Dick et al; Nature Immunology; Volumen 11 Número 7 de julio de 2010
- ^ No es una decisión dividida para la hematopoiesis humana; Kenneth Dorshkind; Nature Immunology Volumen 11 Número 7 de julio de 2010
Imágenes adicionales
Visiones alternativas de linajes
 linaje de células sanguíneas. Para escala, note que los megacariocitos (50-100 μm) son 10 a 15 veces más grandes que un glóbulo rojo típico.
linaje de células sanguíneas. Para escala, note que los megacariocitos (50-100 μm) son 10 a 15 veces más grandes que un glóbulo rojo típico. linaje de células sanguíneas. Basado en la capacidad de auto-renovación.
linaje de células sanguíneas. Basado en la capacidad de auto-renovación. Vista esquemática. linajes bien definidos.
Vista esquemática. linajes bien definidos.- A un lado. Comparando los modelos nuevos y antiguos de linaje.


_diagram_en.svg)
Enlaces externos
- www.copewithcytokines.de Miniportal a la terminología de la linfopoiesis
- Lymphopoiesis en la Biblioteca Nacional de Medicina de EE.UU.
- "Linfopoiesis" en Diccionario médico de Dorland
- Vista general en hematologica.pl