
Intercambio gaseoso (hematosis)

El intercambio de gases o hematosis es el proceso físico por el cual los gases se mueven pasivamente por difusión a través de una superficie. Por ejemplo, esta superficie podría ser la interfaz aire/agua de un cuerpo de agua, la superficie de una burbuja de gas en un líquido, una membrana permeable al gas o una membrana biológica que forma el límite entre un organismo y su entorno extracelular.
Los gases son constantemente consumidos y producidos por reacciones celulares y metabólicas en la mayoría de los seres vivos, por lo que se requiere un sistema eficiente para el intercambio de gases entre, en última instancia, el interior de la(s) célula(s) y el entorno externo. Los organismos pequeños, particularmente los unicelulares, como las bacterias y los protozoos, tienen una alta relación área superficial/volumen. En estas criaturas, la membrana de intercambio de gases suele ser la membrana celular. Algunos pequeños organismos multicelulares, como los gusanos planos, también pueden realizar un intercambio de gases suficiente a través de la piel o la cutícula que rodea sus cuerpos. Sin embargo, en la mayoría de los organismos más grandes, que tienen una relación pequeña entre el área superficial y el volumen, las estructuras especializadas con superficies contorneadas, como branquias, alvéolos pulmonares y mesófilo esponjoso, proporcionan la gran área necesaria para un intercambio gaseoso efectivo. Estas superficies enrevesadas a veces pueden ser internalizadas en el cuerpo del organismo. Es el caso de los alvéolos, que forman la superficie interna del pulmón de los mamíferos, el mesófilo esponjoso, que se encuentra en el interior de las hojas de algunos tipos de plantas, o las branquias de los moluscos que las tienen, que se encuentran en el manto. cavidad.
En los organismos aeróbicos, el intercambio de gases es particularmente importante para la respiración, que implica la absorción de oxígeno ( O
2) y liberación de dióxido de carbono ( CO
2). Por el contrario, en los organismos fotosintéticos oxigénicos, como la mayoría de las plantas terrestres, la absorción de dióxido de carbono y la liberación tanto de oxígeno como de vapor de agua son los principales procesos de intercambio de gases que se producen durante el día. Otros procesos de intercambio de gases son importantes en organismos menos familiares: por ejemplo, el dióxido de carbono, el metano y el hidrógeno se intercambian a través de la membrana celular de las arqueas metanogénicas. En la fijación de nitrógeno por bacterias diazotróficas y la desnitrificación por bacterias heterótrofas (como Paracoccus denitrificans y varias pseudomonas), el gas nitrógeno se intercambia con el medio ambiente, siendo absorbido por las primeras y liberado por las segundas, mientras que los gusanos tubulares gigantes dependen de bacterias para oxidar el sulfuro de hidrógeno extraído de su entorno marino profundo,utilizando oxígeno disuelto en el agua como aceptor de electrones.
La difusión solo tiene lugar con un gradiente de concentración. Los gases fluirán de una concentración alta a una concentración baja. Una alta concentración de oxígeno en los alvéolos y una baja concentración de oxígeno en los capilares hace que el oxígeno se mueva hacia los capilares. Una alta concentración de dióxido de carbono en los capilares y una baja concentración de dióxido de carbono en los alvéolos hace que el dióxido de carbono se mueva hacia los alvéolos.
Principios físicos del intercambio de gases

Difusión y área de superficie
El intercambio de gases se produce como resultado de la difusión a favor de un gradiente de concentración. Las moléculas de gas se mueven de una región en la que están en alta concentración a otra en la que están en baja concentración. La difusión es un proceso pasivo, lo que significa que no se requiere energía para impulsar el transporte, y sigue la Ley de Fick:{displaystyle J=-D{frac {dvarphi}{dx}}}

En relación con un sistema biológico típico, donde dos compartimentos ('interior' y 'exterior') están separados por una barrera de membrana, y donde se permite que un gas se difunda espontáneamente a favor de su gradiente de concentración:
- J es el flujo, la cantidad de gas que se difunde por unidad de área de membrana por unidad de tiempo. Tenga en cuenta que esto ya está escalado para el área de la membrana.
- D es el coeficiente de difusión, que diferirá de gas a gas y de membrana a membrana, según el tamaño de la molécula de gas en cuestión y la naturaleza de la propia membrana (particularmente su viscosidad, temperatura e hidrofobicidad).
- φ es la concentración del gas.
- x es la posición a través del espesor de la membrana.
- d φ /d x es por lo tanto el gradiente de concentración a través de la membrana. Si los dos compartimentos están bien mezclados individualmente, entonces esto se simplifica a la diferencia de concentración del gas entre los compartimentos interior y exterior dividida por el grosor de la membrana.
- El signo negativo indica que la difusión es siempre en la dirección que, con el tiempo, destruirá el gradiente de concentración, es decir, el gas se mueve de una concentración alta a una concentración baja hasta que finalmente los compartimentos interior y exterior alcanzan el equilibrio.
Fig. 1. Ley de Fick para superficies de intercambio de gases
Los gases primero deben disolverse en un líquido para difundirse a través de una membrana, por lo que todos los sistemas biológicos de intercambio de gases requieren un ambiente húmedo. En general, cuanto mayor sea el gradiente de concentración a través de la superficie de intercambio de gases, más rápida será la velocidad de difusión a través de ella. Por el contrario, cuanto más delgada sea la superficie de intercambio de gases (para la misma diferencia de concentración), más rápido se difundirán los gases a través de ella.
En la ecuación anterior, J es el flujo expresado por unidad de área, por lo que aumentar el área no afectará su valor. Sin embargo, un aumento de la superficie disponible, aumentará la cantidad de gas que se puede difundir en un tiempo determinado. Esto se debe a que la cantidad de gas que se difunde por unidad de tiempo (d q /d t ) es el producto de J y el área de la superficie de intercambio de gas, A :{displaystyle {frac{dq}{dt}}=JA}

Los organismos unicelulares, como las bacterias y las amebas, no tienen superficies de intercambio de gases especializadas, porque pueden aprovechar la gran superficie que tienen en relación con su volumen. La cantidad de gas que produce (o requiere) un organismo en un momento dado será aproximadamente proporcional al volumen de su citoplasma. El volumen de un organismo unicelular es muy pequeño, por lo tanto produce (y requiere) una cantidad relativamente pequeña de gas en un tiempo dado. En comparación con este pequeño volumen, el área superficial de su membrana celular es muy grande y adecuada para sus necesidades de intercambio de gases sin modificaciones adicionales. Sin embargo, a medida que un organismo aumenta de tamaño, su área de superficie y su volumen no se escalan de la misma manera. Considere un organismo imaginario que es un cubo de lado L. Su volumen aumenta con el cubo ( L ) de su longitud, pero su superficie externa aumenta solo con el cuadrado ( L ) de su longitud. Esto significa que la superficie externa se vuelve rápidamente inadecuada para las crecientes necesidades de intercambio de gases de un mayor volumen de citoplasma. Además, el grosor de la superficie que deben cruzar los gases ( dx en la Ley de Fick) también puede ser mayor en organismos más grandes: en el caso de un organismo unicelular, una membrana celular típica tiene solo 10 nm de grosor; pero en organismos más grandes como los gusanos redondos (Nematoda), la superficie de intercambio equivalente, la cutícula, es sustancialmente más gruesa a 0,5 μm.
Interacción con los sistemas circulatorios.
En los organismos multicelulares, por lo tanto, los órganos respiratorios especializados, como las branquias o los pulmones, se utilizan a menudo para proporcionar el área de superficie adicional para la tasa requerida de intercambio de gases con el entorno externo. Sin embargo, las distancias entre el intercambiador de gases y los tejidos más profundos suelen ser demasiado grandes para que la difusión satisfaga los requisitos gaseosos de estos tejidos. Por lo tanto, los intercambiadores de gases se acoplan frecuentemente a sistemas circulatorios de distribución de gases, que transportan los gases uniformemente a todos los tejidos corporales independientemente de su distancia del intercambiador de gases.
Algunos organismos multicelulares como los platelmintos (Platyhelminthes) son relativamente grandes pero muy delgados, lo que permite que la superficie externa de su cuerpo actúe como una superficie de intercambio de gases sin necesidad de un órgano de intercambio de gases especializado. Por lo tanto, los platelmintos carecen de branquias o pulmones, y también carecen de un sistema circulatorio. Otros organismos multicelulares como las esponjas (Porifera) tienen un área de superficie inherentemente alta, porque son muy porosos y/o ramificados. Las esponjas no requieren un sistema circulatorio ni órganos especializados de intercambio de gases, porque su estrategia de alimentación implica el bombeo de agua en una sola dirección a través de sus cuerpos porosos utilizando células de collar flageladas. Por lo tanto, cada célula del cuerpo de la esponja está expuesta a un flujo constante de agua fresca oxigenada.
En los organismos que tienen sistemas circulatorios asociados con sus superficies especializadas de intercambio de gases, se utiliza una gran variedad de sistemas para la interacción entre los dos.
En un sistema de flujo a contracorriente, el aire (o, más habitualmente, el agua que contiene aire disuelto) se extrae en la dirección opuesta al flujo de sangre en el intercambiador de gases. Un sistema de contracorriente como este mantiene un fuerte gradiente de concentración a lo largo de la superficie de intercambio de gases (ver diagrama inferior en la Fig. 2). Esta es la situación que se observa en las branquias de los peces y muchas otras criaturas acuáticas. El agua ambiental que contiene gas se atrae unidireccionalmente a través de la superficie de intercambio de gases, con el flujo de sangre en los capilares branquiales debajo fluyendo en la dirección opuesta.Aunque esto teóricamente permite la transferencia casi completa de un gas respiratorio de un lado del intercambiador al otro, en los peces menos del 80% del oxígeno en el agua que fluye por las branquias generalmente se transfiere a la sangre.
Los arreglos alternativos son sistemas de corriente cruzada que se encuentran en las aves. y sistemas de sacos llenos de aire sin salida que se encuentran en los pulmones de los mamíferos. En un sistema de flujo a favor de la corriente, la sangre y el gas (o el fluido que contiene el gas) se mueven en la misma dirección a través del intercambiador de gases. Esto significa que la magnitud del gradiente es variable a lo largo de la superficie de intercambio de gases, y el intercambio eventualmente se detendrá cuando se alcance un equilibrio (ver diagrama superior en la Fig. 2). No se sabe que los sistemas de intercambio de gases de flujo paralelo se utilicen en la naturaleza.
Mamíferos
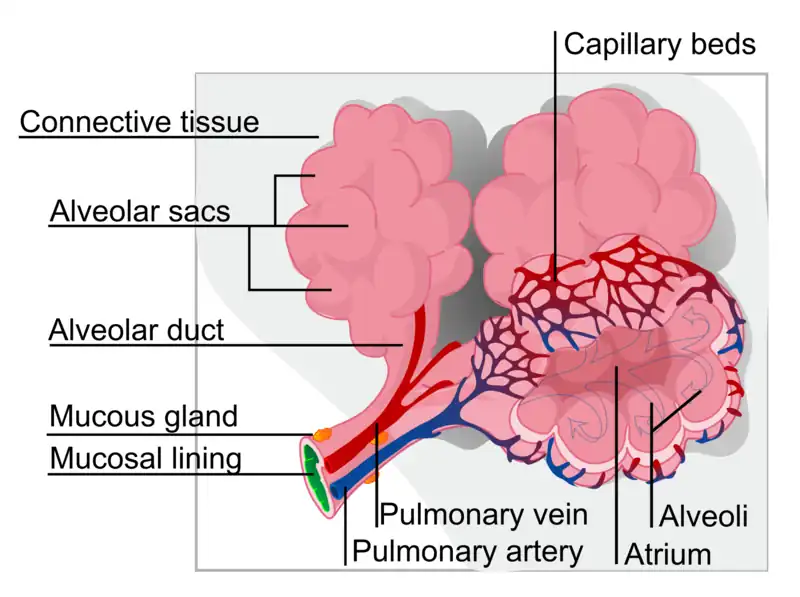
El intercambiador de gases en los mamíferos se internaliza para formar pulmones, como ocurre en la mayoría de los animales terrestres más grandes. El intercambio de gases ocurre en sacos microscópicos llenos de aire sin salida llamados alvéolos, donde una membrana muy delgada (llamada barrera sangre-aire) separa la sangre en los capilares alveolares (en las paredes de los alvéolos) del aire alveolar en los sacos..
Membrana de intercambio
La membrana a través de la cual tiene lugar el intercambio de gases en los alvéolos (es decir, la barrera sangre-aire) es extremadamente delgada (en humanos, en promedio, 2,2 μm de espesor). Está formado por las células epiteliales alveolares, sus membranas basales y las células endoteliales de los capilares pulmonares (fig. 4). La gran superficie de la membrana proviene del plegamiento de la membrana en alrededor de 300 millones de alvéolos, con diámetros de aproximadamente 75-300 μm cada uno. Esto proporciona un área de superficie extremadamente grande (aproximadamente 145 m ) a través de la cual puede ocurrir el intercambio de gases.
Aire alveolar
El aire llega a los alvéolos en pequeñas dosis (llamadas volumen tidal), al inhalar (inhalación) y exhalar (exhalación) a través de las vías respiratorias, un conjunto de tubos relativamente estrechos y moderadamente largos que comienzan en la nariz o la boca y terminan en los alvéolos de los pulmones en el tórax. El aire entra y sale a través del mismo conjunto de tubos, en los que el flujo es en una dirección durante la inhalación y en la dirección opuesta durante la exhalación.
Durante cada inhalación, en reposo, aproximadamente 500 ml de aire fresco entran por la nariz. Se calienta y humedece a medida que fluye a través de la nariz y la faringe. Cuando llega a la tráquea, la temperatura del aire inhalado es de 37 °C y está saturado de vapor de agua. Al llegar a los alvéolos, se diluye y se mezcla completamente con los aproximadamente 2,5 a 3,0 litros de aire que quedaron en los alvéolos después de la última exhalación. Este volumen relativamente grande de aire que está presente de forma semipermanente en los alvéolos durante todo el ciclo respiratorio se conoce como capacidad residual funcional (FRC).
Al comienzo de la inhalación, las vías respiratorias se llenan de aire alveolar inalterado, sobrante de la última exhalación. Este es el volumen del espacio muerto, que suele ser de unos 150 ml. Es el primer aire que vuelve a entrar en los alvéolos durante la inhalación. Solo después de que el aire del espacio muerto haya regresado a los alvéolos, el resto del volumen corriente (500 ml - 150 ml = 350 ml) ingresa a los alvéolos. La entrada de un volumen tan pequeño de aire fresco con cada inhalación, asegura que la composición de la FRC apenas cambie durante el ciclo respiratorio (Fig. 5). La presión parcial alveolar de oxígeno permanece muy cercana a 13-14 kPa (100 mmHg), y la presión parcial de dióxido de carbono varía mínimamente alrededor de 5,3 kPa (40 mmHg) a lo largo del ciclo respiratorio (de inhalación y exhalación).Las presiones parciales correspondientes de oxígeno y dióxido de carbono en el aire ambiente (seco) al nivel del mar son 21 kPa (160 mmHg) y 0,04 kPa (0,3 mmHg) respectivamente.
Este aire alveolar, que constituye el FRC, rodea completamente la sangre en los capilares alveolares (fig. 6). El intercambio de gases en los mamíferos ocurre entre este aire alveolar (que difiere significativamente del aire fresco) y la sangre en los capilares alveolares. Los gases a ambos lados de la membrana de intercambio de gases se equilibran por difusión simple. Esto asegura que las presiones parciales de oxígeno y dióxido de carbono en la sangre que sale de los capilares alveolares y, finalmente, circula por todo el cuerpo, sean las mismas que en el FRC.
La marcada diferencia entre la composición del aire alveolar y la del aire ambiente puede mantenerse porque la capacidad residual funcional está contenida en sacos sin salida conectados al aire exterior por conductos largos y estrechos (las vías respiratorias: nariz, faringe, laringe, tráquea, bronquios y sus ramas y subramas hasta los bronquiolos). Esta anatomía, y el hecho de que los pulmones no se vacían y vuelven a inflar con cada respiración, proporciona a los mamíferos una "atmósfera portátil", cuya composición difiere significativamente del aire ambiental actual.
La composición del aire en el FRC se controla cuidadosamente midiendo las presiones parciales de oxígeno y dióxido de carbono en la sangre arterial. Si la presión de cualquiera de los gases se desvía de lo normal, se provocan reflejos que cambian la frecuencia y la profundidad de la respiración de tal manera que se restablece la normalidad en segundos o minutos.
Circulación pulmonar
Toda la sangre que regresa de los tejidos del cuerpo al lado derecho del corazón fluye a través de los capilares alveolares antes de ser bombeada por todo el cuerpo nuevamente. En su paso a través de los pulmones, la sangre entra en estrecho contacto con el aire alveolar, separado de él por una membrana de difusión muy fina que tiene, en promedio, sólo unos 2 μm de espesor. Por lo tanto, las presiones de los gases en la sangre se equilibrarán rápidamente con las de los alvéolos, asegurando que la sangre arterial que circula por todos los tejidos del cuerpo tenga una tensión de oxígeno de 13-14 kPa (100 mmHg) y una tensión de dióxido de carbono de 5,3 kPa (40 mmHg). Estas presiones arteriales parciales de oxígeno y dióxido de carbono están controladas homeostáticamente. Un aumento en la arteria{displaystyle P_{{mathrm {CO} }_{2}}}

Dado que la sangre que llega a los capilares alveolares tiene un{displaystyle P_{{mathrm {O} }_{2}}}
Solo como resultado de mantener con precisión la composición de los 3 litros de aire alveolar, con cada respiración se libera algo de dióxido de carbono a la atmósfera y se toma algo de oxígeno del aire exterior. Si se ha perdido más dióxido de carbono de lo normal por un breve período de hiperventilación, la respiración se ralentizará o se detendrá hasta que el alveolar{displaystyle P_{{mathrm {CO} }_{2}}}
Si estos homeostáticos están comprometidos, se producirá una acidosis respiratoria o una alcalosis respiratoria. A la larga, estos pueden compensarse mediante ajustes renales en las concentraciones de H y HCO 3 en el plasma; pero dado que esto lleva tiempo, el síndrome de hiperventilación puede, por ejemplo, ocurrir cuando la agitación o la ansiedad hacen que una persona respire rápida y profundamente, expulsando así demasiado CO 2 de la sangre al aire exterior, precipitando una serie de síntomas angustiantes que resultan debido a un pH excesivamente alto de los fluidos extracelulares.
El oxígeno tiene una solubilidad muy baja en agua y, por lo tanto, se transporta en la sangre combinado libremente con la hemoglobina. El oxígeno se mantiene en la hemoglobina por cuatro grupos hemo que contienen hierro ferroso por molécula de hemoglobina. Cuando todos los grupos hemo llevan una molécula de O2 cada uno, se dice que la sangre está "saturada" con oxígeno, y ningún aumento adicional en la presión parcial de oxígeno aumentará significativamente la concentración de oxígeno en la sangre. La mayor parte del dióxido de carbono en la sangre se transporta como iones HCO 3 en el plasma. Sin embargo, la conversión de CO 2 disuelto en HCO 3(a través de la adición de agua) es demasiado lento para la velocidad a la que circula la sangre a través de los tejidos, por un lado, y los capilares alveolares, por el otro. Por lo tanto, la reacción es catalizada por la anhidrasa carbónica, una enzima dentro de los glóbulos rojos. La reacción puede ir en cualquier dirección dependiendo de la presión parcial predominante de dióxido de carbono. Una pequeña cantidad de dióxido de carbono se transporta en la porción proteica de las moléculas de hemoglobina como grupos carbamino. La concentración total de dióxido de carbono (en forma de iones bicarbonato, CO 2 disuelto y grupos carbamino) en la sangre arterial (es decir, después de que se haya equilibrado con el aire alveolar) es de aproximadamente 26 mM (o 58 ml/100 ml).en comparación con la concentración de oxígeno en sangre arterial saturada de aproximadamente 9 mM (o 20 ml/100 ml de sangre).
Otros vertebrados
Pescado
El contenido de oxígeno disuelto en el agua dulce es de aproximadamente 8 a 10 mililitros por litro en comparación con el del aire, que es de 210 mililitros por litro. El agua es 800 veces más densa que el aire y 100 veces más viscosa. Por tanto, el oxígeno tiene una velocidad de difusión en el aire 10.000 veces mayor que en el agua. Por lo tanto, el uso de pulmones en forma de saco para eliminar el oxígeno del agua no sería lo suficientemente eficiente para sustentar la vida. En lugar de usar los pulmones, el intercambio gaseoso tiene lugar a través de la superficie de branquias altamente vascularizadas. Las branquias son órganos especializados que contienen filamentos, que luego se dividen en láminas. Las laminillas contienen capilares que proporcionan una gran superficie y cortas distancias de difusión, ya que sus paredes son extremadamente delgadas.Los rastrillos branquiales se encuentran dentro del sistema de intercambio para filtrar los alimentos y mantener limpias las branquias.
Las branquias utilizan un sistema de flujo a contracorriente que aumenta la eficiencia de la absorción de oxígeno (y la pérdida de gases residuales). El agua oxigenada se aspira a través de la boca y pasa por las branquias en una dirección, mientras que la sangre fluye a través de las láminas en la dirección opuesta. Esta contracorriente mantiene fuertes gradientes de concentración a lo largo de cada capilar (consulte el diagrama en la sección anterior "Interacción con los sistemas circulatorios"). El oxígeno es capaz de difundirse continuamente por su gradiente hacia la sangre, y el dióxido de carbono por su gradiente hacia el agua.El agua desoxigenada eventualmente pasará a través del opérculo (cubierta branquial). Aunque los sistemas de intercambio de contracorriente teóricamente permiten una transferencia casi completa de un gas respiratorio de un lado del intercambiador al otro, en los peces menos del 80% del oxígeno en el agua que fluye por las branquias generalmente se transfiere a la sangre.
Anfibios
Los anfibios tienen tres órganos principales involucrados en el intercambio de gases: los pulmones, la piel y las branquias, que pueden usarse solos o en una variedad de combinaciones diferentes. La importancia relativa de estas estructuras difiere según la edad, el ambiente y la especie del anfibio. La piel de los anfibios y sus larvas está muy vascularizada, lo que da lugar a un intercambio de gases relativamente eficaz cuando la piel está húmeda. Las larvas de los anfibios, como la etapa de renacuajo de las ranas antes de la metamorfosis, también tienen branquias externas. Las branquias se absorben en el cuerpo durante la metamorfosis, después de lo cual los pulmones se harán cargo. Los pulmones suelen ser más simples que en los otros vertebrados terrestres, con pocos tabiques internos y alvéolos más grandes; sin embargo, los sapos, que pasan más tiempo en tierra, tienen una superficie alveolar más grande con pulmones más desarrollados.El piso inferior de la boca se mueve en forma de "bombeo", lo que se puede observar a simple vista.
Reptiles
Todos los reptiles respiran usando pulmones. En los escamatos (los lagartos y las serpientes), la ventilación es impulsada por la musculatura axial, pero esta musculatura también se usa durante el movimiento, por lo que algunos escamatos dependen del bombeo bucal para mantener la eficiencia del intercambio de gases.
Debido a la rigidez de los caparazones de tortuga y tortuga, es difícil una expansión y contracción significativas del tórax. Las tortugas terrestres y terrestres dependen de las capas musculares adheridas a sus caparazones, que envuelven sus pulmones para llenarlos y vaciarlos. Algunas tortugas acuáticas también pueden bombear agua en una boca o cloaca altamente vascularizada para lograr el intercambio de gases.
Los cocodrilos tienen una estructura similar al diafragma de los mamíferos, el diafragmaticus, pero este músculo ayuda a crear un flujo de aire unidireccional a través de los pulmones en lugar de un flujo de marea: esto es más similar al flujo de aire que se ve en las aves que al que se ve en los mamíferos. Durante la inhalación, el diafragma hace retroceder al hígado, inflando los pulmones en el espacio que esto crea. El aire fluye hacia los pulmones desde los bronquios durante la inhalación, pero durante la exhalación, el aire sale de los pulmones hacia los bronquios por una ruta diferente: este movimiento de gas en una sola dirección se logra mediante válvulas aerodinámicas en las vías respiratorias.
Aves
Las aves tienen pulmones pero no diafragma. Se basan principalmente en sacos de aire para la ventilación. Estos sacos de aire no juegan un papel directo en el intercambio de gases, pero ayudan a mover el aire unidireccionalmente a través de las superficies de intercambio de gases en los pulmones. Durante la inhalación, el aire fresco se toma de la tráquea hacia los sacos de aire posteriores y hacia los parabronquios que conducen desde los sacos de aire posteriores al pulmón. El aire que ingresa a los pulmones se une al aire que ya está en los pulmones y es arrastrado a través del intercambiador de gases hacia los sacos de aire anteriores. Durante la exhalación, los alvéolos posteriores fuerzan el aire hacia los mismos parabronquios de los pulmones, fluyendo en la misma dirección que durante la inhalación, lo que permite un intercambio continuo de gases independientemente del ciclo respiratorio.La broncoconstricción selectiva en los diversos puntos de ramificación bronquial asegura que el aire no fluya y fluya a través de los bronquios durante la inhalación y la exhalación, como ocurre en los mamíferos, sino que siga los caminos descritos anteriormente.
El flujo de aire unidireccional a través de los parabronquios intercambia gases respiratorios con un flujo sanguíneo cruzado (Fig. 9). La presión parcial de O 2 ({displaystyle P_{{mathrm {O} }_{2}}}
Plantas
El intercambio de gases en las plantas está dominado por las funciones del dióxido de carbono, el oxígeno y el vapor de agua. CO
2es la única fuente de carbono para el crecimiento autótrofo por fotosíntesis, y cuando una planta realiza activamente la fotosíntesis a la luz, absorberá dióxido de carbono y perderá vapor de agua y oxígeno. Por la noche, las plantas respiran y el intercambio de gases se invierte parcialmente: todavía se pierde vapor de agua (pero en menor medida), pero ahora se absorbe oxígeno y se libera dióxido de carbono.
El intercambio de gases de las plantas ocurre principalmente a través de las hojas. El intercambio de gases entre una hoja y la atmósfera ocurre simultáneamente a través de dos vías: 1) células epidérmicas y ceras cuticulares (generalmente denominadas "cutícula") que siempre están presentes en la superficie de cada hoja, y 2) estomas, que normalmente controlan la mayoría de las intercambio. Los gases entran en el tejido fotosintético de la hoja a través de la disolución en la superficie húmeda de la empalizada y las células del mesófilo esponjoso. Las células del mesófilo esponjoso están empaquetadas de forma suelta, lo que permite una mayor área de superficie y, en consecuencia, una mayor tasa de intercambio de gases. La absorción de dióxido de carbono necesariamente resulta en alguna pérdida de vapor de agua, porque ambas moléculas entran y salen por los mismos estomas, por lo que las plantas experimentan un dilema de intercambio de gases: ganar suficienteCO
2sin perder demasiada agua. Por lo tanto, la cutícula cerosa de la epidermis de la hoja minimiza la pérdida de agua de otras partes de la hoja. El tamaño de un estoma está regulado por la apertura y el cierre de sus dos células de guarda: la turgencia de estas células determina el estado de la apertura del estoma, y este a su vez está regulado por el estrés hídrico. Las plantas que muestran un metabolismo ácido de las crasuláceas son xerófitas tolerantes a la sequía y realizan casi todo su intercambio de gases durante la noche, porque solo durante la noche estas plantas abren sus estomas. Al abrir los estomas solo por la noche, se minimiza la pérdida de vapor de agua asociada con la absorción de dióxido de carbono. Sin embargo, esto tiene el costo de un crecimiento lento: la planta tiene que almacenar el dióxido de carbono en forma de ácido málico para usarlo durante el día, y no puede almacenar cantidades ilimitadas.
Las mediciones de intercambio de gases son herramientas importantes en la ciencia de las plantas: esto generalmente implica sellar la planta (o parte de una planta) en una cámara y medir los cambios en la concentración de dióxido de carbono y vapor de agua con un analizador de gases infrarrojo. Si las condiciones ambientales (humedad, CO
2concentración, luz y temperatura) están totalmente controlados, las mediciones de CO
2la absorción y la liberación de agua revelan información importante sobre el CO
2tasas de asimilación y transpiración. El CO intercelular
2concentración revela información importante sobre la condición fotosintética de las plantas. Se pueden usar métodos más simples en circunstancias específicas: el indicador de hidrogenocarbonato se puede usar para monitorear el consumo de CO
2en una solución que contiene una sola hoja de planta a diferentes niveles de intensidad de luz, y la generación de oxígeno por la alga marina Elodea se puede medir simplemente recolectando el gas en un tubo de ensayo sumergido que contiene una pequeña parte de la planta.
Invertebrados
El mecanismo de intercambio de gases en los invertebrados depende de su tamaño, estrategia de alimentación y hábitat (acuático o terrestre).
Las esponjas (Porifera) son criaturas sésiles, lo que significa que no pueden moverse por sí mismas y normalmente permanecen adheridas a su sustrato. Obtienen nutrientes a través del flujo de agua a través de sus células e intercambian gases por simple difusión a través de sus membranas celulares. Los poros llamados ostia atraen agua hacia la esponja y el agua circula posteriormente a través de la esponja por células llamadas coanocitos que tienen estructuras similares a cabellos que mueven el agua a través de la esponja.
Los cnidarios incluyen corales, anémonas de mar, medusas e hidras. Estos animales siempre se encuentran en ambientes acuáticos, que van desde agua dulce hasta agua salada. No tienen ningún órgano respiratorio dedicado; en cambio, cada célula de su cuerpo puede absorber oxígeno del agua circundante y liberarle gases residuales. Una desventaja clave de esta característica es que los cnidarios pueden morir en ambientes donde el agua está estancada, ya que agotan el suministro de oxígeno del agua. Los corales a menudo forman simbiosis con otros organismos, en particular con los dinoflagelados fotosintéticos. En esta simbiosis, el coral brinda refugio y el otro organismo proporciona nutrientes al coral, incluido el oxígeno.
Los gusanos redondos (Nematoda), los platelmintos (Platyhelminthes) y muchos otros pequeños animales invertebrados que viven en hábitats acuáticos o húmedos no tienen una superficie dedicada al intercambio de gases ni un sistema circulatorio. En su lugar, se basan en la difusión de CO
2y O
2directamente a través de su cutícula. La cutícula es la capa más externa semipermeable de sus cuerpos.
Otros invertebrados acuáticos, como la mayoría de los moluscos (Mollusca) y los crustáceos más grandes (Crustacea), como las langostas, tienen branquias análogas a las de los peces, que funcionan de manera similar.
A diferencia de los grupos de invertebrados mencionados hasta ahora, los insectos suelen ser terrestres e intercambian gases a través de una superficie húmeda en contacto directo con la atmósfera, en lugar de estar en contacto con el agua circundante. El exoesqueleto del insecto es impermeable a los gases, incluido el vapor de agua, por lo que tienen un sistema de intercambio de gases más especializado, que requiere que los gases se transporten directamente a los tejidos a través de una compleja red de tubos. Este sistema respiratorio está separado de su sistema circulatorio. Los gases entran y salen del cuerpo a través de aberturas llamadas espiráculos, ubicadas lateralmente a lo largo del tórax y el abdomen. Al igual que las plantas, los insectos pueden controlar la apertura y el cierre de estos espiráculos, pero en lugar de depender de la presión de la turgencia, dependen de las contracciones musculares.Estas contracciones hacen que el abdomen de un insecto se bombee hacia adentro y hacia afuera. Los espiráculos están conectados a tubos llamados tráqueas, que se ramifican repetidamente y se ramifican en el cuerpo del insecto. Estas ramas terminan en células traqueales especializadas que proporcionan una superficie delgada y húmeda para un intercambio de gases eficiente, directamente con las células.
El otro grupo principal de artrópodos terrestres, los arácnidos (arañas, escorpiones, ácaros y sus parientes) suelen realizar el intercambio de gases con un pulmón de libro.
Resumen de los principales sistemas de intercambio de gases
| Área de superficie | Distancia de difusión | Mantenimiento del gradiente de concentración | órganos respiratorios | |
|---|---|---|---|---|
| Humano | Alvéolos totales = 70–100 m | Alvéolo y capilar (dos células) | Flujo sanguíneo constante en los capilares; respiración | Pulmones |
| Pescado | Muchas laminillas y filamentos por branquia | Por lo general, una celda | Flujo a contracorriente | Branquias |
| Insectos | Célula traqueal especializada | una celda | Bombeo bucal | espiráculos |
| esponjas | poros de ostia | una celda | Movimiento de agua | Ninguna |
| gusanos planos | forma de cuerpo plano | Por lo general, una celda | Flujo a contracorriente | Ninguna |
| cnidarios | brazos orales | Por lo general, una celda | Movimiento de agua | Ninguna |
| Reptiles | Muchas laminillas y filamentos por branquia | Alvéolo y capilar (dos células) | Flujo a contracorriente | Pulmones |
| anfibios | Muchas laminillas y filamentos por branquia | Alvéolo y capilar (dos células) o una célula | Flujo a contracorriente | Pulmones, piel y branquias |
| Plantas | Alta densidad de estomas; espacios de aire dentro de la hoja | una celda | Flujo de aire constante | estomas |
Contenido relacionado
Pitón reticulada
Estela (planta)
Hemeritrina