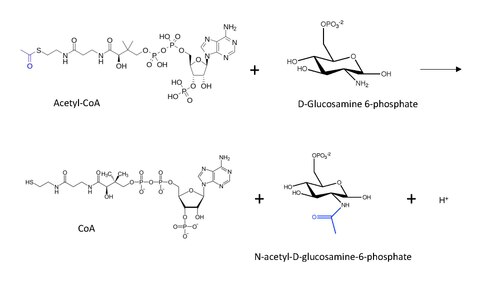
En enzimología, la glucosamina-fosfato N-acetiltransferasa (GNA) (EC 2.3.1.4) es una enzima que cataliza la transferencia de un grupo acetilo del acetil-CoA a la amina primaria en la glucosamida-6-fosfato, generando un CoA libre y N-acetil-D-glucosamina-6-fosfato.Esta enzima pertenece a la familia de las transferasas, un grupo de enzimas que transfiere un grupo funcional muy específico, en este caso el acetilo, de un donante a un receptor. Específicamente, esta enzima puede caracterizarse como parte de la familia de las aciltransferasas, ya que implica la transferencia de un grupo acilo general con un metilo como sustituyente.
Nomenclature
El nombre sistemático de esta clase de enzimas es acetil-CoA:D-glucosamina-6-fosfato N-acetiltransferasa. Otros nombres comunes incluyen fosfoglucosamina transacetilasa, fosfoglucosamina acetilasa, glucosamina-6-fosfato acetilasa, D-glucosamina-6-P N-acetiltransferasa, aminodesoxiglucosafosfato acetiltransferasa, glucosamina 6-fosfato acetilasa, glucosamina 6-fosfato N-acetiltransferasa, N-acetilglucosamina-6-fosfato sintasa, fosfoglucosamina N-acetilasa, glucosamina-fosfato N-acetiltransferasa y glucosamina-6-fosfato N-acetiltransferasa.
Función
Esta enzima forma parte de la vía de biosíntesis de la hexosamina (HBP), una de las vías de procesamiento de la glucosa en el metabolismo general. Esta vía comparte los dos pasos iniciales con la glucólisis y difiere solo una pequeña porción del flujo de glucosa de esta vía glucolítica más tradicional. Por lo tanto, se ve favorecida cuando existe una regulación por retroalimentación negativa de la glucólisis, como en el caso de grandes cantidades de ácidos grasos libres. El producto final de esta vía es la UDP-N-acetilglucosamina, que participa en la modificación de moléculas complejas como los glucolípidos, los proteoglicanos y las glucoproteínas. Este producto final actúa como transportador de la N-acetilglucosamina, unidad monomérica de la quitina, un polímero estructural que compone las conchas de crustáceos e insectos, así como la pared celular de los hongos. Además, la N-acetilglucosamina también es una unidad del polímero de peptidoglicano que compone la pared celular bacteriana junto con el disacárido N-acetilmurámico.Figura 1: Como se muestra en la imagen, hay dos caminos posibles para producir UDP-N-Acetyl-D-Glucosamida de D-Fructose-6-fosfato. En eucariotas, el GNA está presente y la acetilación ocurre antes de la transferencia de fosfato de 6-carbono a 1-carbono. En prokaryotes, el orden se invierte, y el sustrato de acilación es más bien D-Glucosamida-1-fosfato, y una sola enzima cataliza tanto la transferencia de carbono como la unión UTP, generando directamente UDP-GlcNAc (UDP-N-Acetyl-D-Glucosamide). Imagen reproducida.Más específicamente, la enzima GNA cataliza el cuarto paso de la vía HBP en eucariotas, promoviendo la transferencia de carbono del acetil-CoA al otro sustrato, la D-glucosamina-6-fosfato, que finalmente producirá UDP-N-acetilglucosamina. Este es un paso químico pequeño, pero importante, crucial para las propiedades de los subproductos de esta vía metabólica. La acetilación se lleva a cabo hasta el producto final de la vía de la hexamina y es muy característica de los polímeros formados con N-acetilglucosamina. Por ejemplo, constituye una de las principales diferencias en la estructura molecular de la quitina y la celulosa, y explica muchas de las propiedades físicas y químicas de estos polímeros. En el caso de la quitina, por ejemplo, estudios computacionales han demostrado que la acilación contribuye a la formación de enlaces de hidrógeno que estabilizan la estructura cristalina de este polímero, proporcionando una mayor resistencia a la fractura.Sin embargo, en el metabolismo procariota, la vía de biosíntesis de hexosamina sigue una etapa de reacción diferente, en la que una enzima distinta actúa sobre los mismos sustratos característicos (Figura 1). En procariotas, la transferencia de fosfato del carbono 6 al carbono 1 ocurre antes de la acilación, de modo que el sustrato de la reacción de adición de carbono es la glucosamina-1-fosfato en lugar de la D-glucosamina-6-fosfato. En esta ocasión, la enzima responsable de la acetilación es la proteína bifuncional glmU (N-acetilglucosamina-1-fosfato uridiltransferasa), que también cataliza la adición de UDP al grupo fosfato de la N-acetil-D-glucosamina-1-fosfato.En humanos, la glucosamina-fosfato N-acetiltransferasa es un dímero con dos subunidades idénticas y está codificada en el gen GNPNAT (símbolo HGNC). Más específicamente, la enzima se expresa con fuerza en los tejidos del hígado, el estómago y el tracto gastrointestinal, y dentro de la célula, se localiza en los endosomas y en el aparato de Golgi (según anotación manual).
Mecanismo
Esquema que muestra la reacción catalizada por Glucosamina-6-fosfato N-acetilltransferase, mostrando acyl grupo en azul transferido de Acetyl CoA al hexosamina Glucosamina-6-fosfato.La estructura molecular de la reacción catalizada por GNA se muestra a continuación, con el grupo acetilo transferido en azul.El mecanismo general de reacción postulado para la acetilación del extremo N de la proteína (inspirado en el mecanismo de acetilación de la lisina) con acetil-CoA implica un ataque nucleofílico del grupo amino (en este caso, de D-glucosamina-6-fosfato) sobre el carbonilo terminal en la transferencia de carbono, lo que conduce a la formación de un intermedio tetraédrico carbonado. La reacción procede con la restauración del carbonilo mediante la eliminación del CoA como grupo saliente, de modo que ahora el grupo acetilo está conectado al grupo amino en el otro sustrato.Específicamente para estos catalizadores de N-acetiltransferasa, estudios con la enzima GNA de S. cerevisiae han demostrado que algunos aminoácidos específicos contribuyen a la unión del sustrato, al aumento de la nucleofilicidad del grupo amino y, finalmente, a la catálisis, lo que respalda el mecanismo postulado descrito anteriormente. Glu98, Asp99 e Ile100 polarizan el enlace carbonilo en el acetil-CoA, lo que aumenta la electrofilicidad del carbono y estabiliza el intermedio tetraédrico del carbono. Tyr143 es responsable de estabilizar el anión tiolato, favoreciendo la S-CoA como grupo saliente del carbono tetraédrico. Finalmente, Asp134 mejora la nucleofilicidad del grupo amino en la D-glucosamida-6-fosfato al ceder densidad electrónica al átomo de nitrógeno. En un organismo diferente, C. albicans, se encontró que un conjunto similar de aminoácidos era esencial para la actividad catalítica: el sistema Glu88-Asp-89-Ile90, Asp125 y Tyr133, respectivamente.
Estructura
Figura 3: Estructura de cristal del ARN unido a sustratos. Estructura de la Base de Datos de ProteínaA finales de 2019, se habían resuelto 13 estructuras para esta clase de enzimas en diferentes especies, con los códigos de acceso PDB 1I12 (Saccharomyces cerevisiae), 1I1D (Saccharomyces cerevisiae), 1I21 (Saccharomyces cerevisiae), 2HUZ (Homo sapiens), 2O28 (Homo sapiens), 4AG7 (Caenorhabditis elegans), entre otros.La Figura 3 muestra la estructura cristalina propuesta de GNA en humanos, con cada subunidad catalítica en un color diferente. El acetil-CoA unido a la enzima se muestra en rosa claro, y el producto aún unido al sitio catalítico se muestra en morado. El grupo acetilo transferido en el producto N-acetil-D-glucosamina-6-fosfato (morado) se muestra en amarillo. Esta propuesta de estructura tridimensional de la proteína muestra que las partes específicas de los sustratos involucrados en esta reacción (el extremo terminal de la porción lineal del acetil-CoA y el grupo nitrógeno unido al anillo de glucosamina) están muy próximas.
Referencias
^Kato N, Mueller CR, Wessely V, Lan Q, Christensen BM (junio de 2005). "Mosquito glucosamine-6-fosfato N-acetilransferase: cDNA, estructura genética y kinetics enzima". Bioquímica de insectos y biología molecular. 35 (6): 637 –46. Bibcode:2005IBMB...35..637K. doi:10.1016/j.ibmb.2005.02.005. PMID 15857769.
^Schwarzer M, Doenst T (2016). Guía del Científico para el Metabolismo Cardiaco. Prensa Académica. pp. 39 –55. ISBN 9780128023945.
^Kim YH, Nakayama T, Nayak J (enero 2018). "Glycolysis y la vía biosintética de Hexosamina como objetivos de la inflamación de la vía aérea superior e inferior". Alergia, Asma & Inmunología Research. 10 1): 6 –11. doi:10.4168/aair.2018.10.1.6. 5705485. PMID 29178672.
^Cohen E (octubre de 2001). "Sintesis e inhibición de la mentina: un revisit". Pest Management Science. 57 (10): 946 –50. doi:10.1002/ps.363. PMID 11695188.
^Meroueh SO, Bencze KZ, Hesek D, Lee M, Fisher JF, Stemmler TL, Mobashery S (marzo de 2006). "La estructura tridimensional de la pared celular bacteriana peptidoglycan". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América. 103 (12): 4404 –9. Bibcode:2006PNAS..103.4404M. doi:10.1073/pnas.0510182103. PMC 1450184. PMID 16537437.
^ a bRiegler H, Herter T, Grishkovskaya I, Lude A, Ryngajllo M, Bolger ME, Essigmann B, Usadel B (abril de 2012). "Crystal structure and functional characterization of a glucosamine-6-phosphate N-acetyltransferase from Arabidopsis thaliana". The Biochemical Journal. 443 2): 427 –37. doi:10.1042/BJ20112071. PMID 22329777.
^"Fig. 1 Estructura química de celulosa y chitín". ResearchGate. Retrieved 2019-03-15.
^Cui J, Yu Z, Lau D (enero de 2016). "Efecto del grupo Acetyl sobre propiedades mecánicas de Chitin/Chitosan Nanocrystal: Un estudio dinámico molecular". International Journal of Molecular Sciences. 17 1): 61. doi:10.3390/ijms17010061. PMC 4730306. PMID 26742033.
^Vithani N, Bais V, Prakash B (junio de 2014). "GlmU (N-acetylglucosamina-1-fosfato uridyltransferase) atado a tres iones de magnesio y ATP en el sitio activo". Acta Crystallographica Section F. 70 (Pt 6): 703 –8. doi:10.1107/S2053230X14008279. PMC 4051520. PMID 24915076.
^Wang J, Liu X, Liang YH, Li LF, Su XD (septiembre de 2008). "Acceptor substrate binding revealed by cristal structure of human glucosamine-6-phosphate N-acetyltransferase 1". Cartas FEBS. 582 (20): 2973 –8. Bibcode:2008FEBSL.582.2973W. doi:10.1016/j.febslet.2008.07.040. PMID 18675810. S2CID 5131540.
^ a b"GNPNAT1 - Glucosamina 6 fosfato N-acetyltransferase - Homo sapiens (Human) - GNPNAT1 gen < protein". www.uniprot.org. Retrieved 2019-03-15.
^Lim S, Smith KR, Lim ST, Tian R, Lu J, Tan M (2016-04-14). "Regulación de funciones mitocondriales por fosforilación proteica y defosforilación". Cell " Bioscience. 625. doi:10.1186/s13578-016-0089-3. PMC 4832502. PMID 27087918.
^Mio T, Yamada-Okabe T, Arisawa M, Yamada-Okabe H (enero de 1999). "Saccharomyces cerevisiae GNA1, un gen esencial encodificando una novela acetyltransferase involucrada en la síntesis UDP-N-acetylglucosamine". El Diario de Química Biológica. 274 1): 424 –9. doi:10.1074/jbc.274.1.424. PMID 9867860.
^Milewski S, Gabriel I, Olchowy J (enero 2006). "Enzimas de biosíntesis UDP-GlcNAc en levadura". Levadura. 23 1): 1 –14. doi:10.1002/yea.1337. PMID 16408321. S2CID 39940329.
^PDB: 2O28; Plotnikov AN, Bochkarev A, Edwards AM, Arrowsmith CH, Sundstrom M, Weigelt J, et al. "Crystal Structure of Glucosamine-Phosphate N-Acetyltransferase 1". Worldwide Protein Data Bank. doi:10.2210/pdb2o28/pdb.
^Peneff C, Mengin-Lecreulx D, Bourne Y (mayo de 2001). "Las estructuras cristalinas de Apo y complejos Saccharomyces cerevisiae GNA1 arrojan luz sobre el mecanismo catalítico de un amino-aztyltransferase de azúcar". El Diario de Química Biológica. 276 (19): 16328 –34. doi:10.1074/jbc.M009988200. PMID 11278591.