
Glucólisis
La glucólisis es la vía metabólica que convierte la glucosa (C 6 H 12 O 6), en ácido pirúvico (CH 3 COCO 2 H). La energía libre liberada en este proceso se utiliza para formar moléculas de alta energía, trifosfato de adenosina (ATP) y dinucleótido de nicotinamida y adenina reducido (NADH). La glucólisis es una secuencia de diez reacciones catalizadas por enzimas.
La glucólisis es una vía metabólica que no requiere oxígeno. La amplia presencia de glucólisis en otras especies indica que se trata de una vía metabólica antigua. De hecho, las reacciones que componen la glucólisis y su vía paralela, la vía de las pentosas fosfato, ocurren en las condiciones libres de oxígeno de los océanos del Arcaico, también en ausencia de enzimas, catalizadas por metales.
En la mayoría de los organismos, la glucólisis ocurre en la parte líquida de las células, el citosol. El tipo más común de glucólisis es la vía Embden-Meyerhof-Parnas (EMP), que fue descubierta por Gustav Embden, Otto Meyerhof y Jakub Karol Parnas. La glucólisis también se refiere a otras vías, como la vía Entner-Doudoroff y varias vías heterofermentativas y homofermentativas. Sin embargo, la discusión aquí se limitará a la vía Embden-Meyerhof-Parnas.
La vía de la glucólisis se puede separar en dos fases:
- Fase de inversión: en la que se consume ATP
- Fase de rendimiento: en la que se produce más ATP del que se consume originalmente
Visión de conjunto
La reacción general de la glucólisis es:
d -Glucosa

+ 2 [NAD]+ 2 [ADP]+ 2 [P]
yo
2 × piruvato
2 × 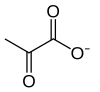 + 2 [NADH]+ 2 H+ 2 [ATP]+ 2 H
+ 2 [NADH]+ 2 H+ 2 [ATP]+ 2 H
2 O
El uso de símbolos en esta ecuación hace que parezca desequilibrada con respecto a los átomos de oxígeno, los átomos de hidrógeno y las cargas. El equilibrio atómico lo mantienen los dos grupos fosfato (P i):
- Cada uno existe en forma de anión de fosfato de hidrógeno (HPO 4), disociándose para contribuir con 2 H en general
- Cada uno libera un átomo de oxígeno cuando se une a una molécula de difosfato de adenosina (ADP), contribuyendo con 2 O en total.
Los cargos se equilibran por la diferencia entre ADP y ATP. En el ambiente celular, los tres grupos hidroxilo de ADP se disocian en −O y H, dando ADP, y este ion tiende a existir en un enlace iónico con Mg, dando ADPMg. El ATP se comporta de manera idéntica excepto que tiene cuatro grupos hidroxilo, dando ATPMg. Cuando estas diferencias junto con las cargas verdaderas en los dos grupos fosfato se consideran juntas, las cargas netas de -4 en cada lado están balanceadas.
Para fermentaciones simples, el metabolismo de una molécula de glucosa a dos moléculas de piruvato tiene un rendimiento neto de dos moléculas de ATP. La mayoría de las células llevarán a cabo más reacciones para "devolver" el NAD usado y producir un producto final de etanol o ácido láctico. Muchas bacterias utilizan compuestos inorgánicos como aceptores de hidrógeno para regenerar el NAD.
Las células que realizan la respiración aeróbica sintetizan mucho más ATP, pero no como parte de la glucólisis. Estas reacciones aeróbicas adicionales utilizan piruvato y NADH + H de la glucólisis. La respiración aeróbica eucariótica produce aproximadamente 34 moléculas adicionales de ATP por cada molécula de glucosa, sin embargo, la mayoría de estas se producen mediante un mecanismo muy diferente de la fosforilación a nivel de sustrato en la glucólisis.
La menor producción de energía, por glucosa, de la respiración anaeróbica en relación con la respiración aeróbica, da como resultado un mayor flujo a través de la vía en condiciones hipóxicas (poco oxígeno), a menos que se encuentren fuentes alternativas de sustratos oxidables anaeróbicamente, como los ácidos grasos.
Historia
La vía de la glucólisis, tal como se la conoce hoy en día, tardó casi 100 años en dilucidarse por completo. Se requirieron los resultados combinados de muchos experimentos más pequeños para comprender la vía en su conjunto.
Los primeros pasos para comprender la glucólisis comenzaron en el siglo XIX con la industria del vino. Por razones económicas, la industria vitivinícola francesa buscó investigar por qué el vino a veces se volvía desagradable, en lugar de fermentar hasta convertirse en alcohol. El científico francés Louis Pasteur investigó este tema durante la década de 1850, y los resultados de sus experimentos iniciaron el largo camino para dilucidar el camino de la glucólisis. Sus experimentos demostraron que la fermentación ocurre por la acción de microorganismos vivos, levaduras, y que el consumo de glucosa de la levadura disminuyó en condiciones aeróbicas de fermentación, en comparación con las condiciones anaeróbicas (el efecto Pasteur).
Los experimentos de fermentación no celular de Eduard Buchner durante la década de 1890 proporcionaron información sobre los pasos que componen la glucólisis. Buchner demostró que la conversión de glucosa en etanol era posible utilizando un extracto no vivo de levadura, debido a la acción de las enzimas del extracto. Este experimento no solo revolucionó la bioquímica, sino que también permitió a los científicos posteriores analizar esta vía en un entorno de laboratorio más controlado. En una serie de experimentos (1905-1911), los científicos Arthur Harden y William Young descubrieron más fragmentos de glucólisis. Descubrieron los efectos reguladores del ATP sobre el consumo de glucosa durante la fermentación alcohólica. También arrojan luz sobre el papel de un compuesto como intermediario de la glucólisis: la fructosa 1,6-bisfosfato.
La elucidación de la fructosa 1,6-bisfosfato se logró midiendo los niveles de CO2 cuando el jugo de levadura se incubó con glucosa. La producción de CO 2 aumentó rápidamente y luego se desaceleró. Harden y Young notaron que este proceso se reiniciaría si se agregara un fosfato inorgánico (Pi) a la mezcla. Harden y Young dedujeron que este proceso producía ésteres de fosfato orgánico, y experimentos posteriores les permitieron extraer fructosa difosfato (F-1,6-DP).
Arthur Harden y William Young junto con Nick Sheppard determinaron, en un segundo experimento, que una fracción subcelular de alto peso molecular sensible al calor (las enzimas) y una fracción de citoplasma de bajo peso molecular insensible al calor (ADP, ATP y NAD y otros cofactores) se requieren juntos para que proceda la fermentación. Este experimento comenzó observando que el jugo de levadura dializado (purificado) no podía fermentar o incluso crear un fosfato de azúcar. Esta mezcla se rescató con la adición de extracto de levadura sin dializar que se había hervido. Hervir el extracto de levadura inactiva todas las proteínas (ya que las desnaturaliza). La capacidad del extracto hervido más el jugo dializado para completar la fermentación sugiere que los cofactores no tenían un carácter proteico.
En la década de 1920, Otto Meyerhof pudo unir algunas de las muchas piezas individuales de glucólisis descubiertas por Buchner, Harden y Young. Meyerhof y su equipo pudieron extraer diferentes enzimas glicolíticas del tejido muscular y combinarlas para crear artificialmente la vía del glucógeno al ácido láctico.
En un artículo, Meyerhof y la científica Renate Junowicz-Kockolaty investigaron la reacción que divide la fructosa 1,6-difosfato en dos triosas fosfatos. El trabajo anterior propuso que la división se produjo a través de 1,3-difosfogliceraldehído más una enzima oxidante y cosomasa. Meyerhoff y Junowicz encontraron que la constante de equilibrio para la reacción de isomerasa y aldosa no se ve afectada por fosfatos inorgánicos o cualquier otra cosomasa o enzimas oxidantes. Además, eliminaron el difosfogliceraldehído como posible intermediario en la glucólisis.
Con todas estas piezas disponibles en la década de 1930, Gustav Embden propuso un esquema detallado paso a paso de esa vía que ahora conocemos como glucólisis. Las mayores dificultades para determinar las complejidades de la ruta se debieron a la vida útil muy corta y las bajas concentraciones en estado estacionario de los intermedios de las reacciones glucolíticas rápidas. En la década de 1940, Meyerhof, Embden y muchos otros bioquímicos finalmente completaron el rompecabezas de la glucólisis. La comprensión de la vía aislada se ha ampliado en las décadas posteriores, para incluir más detalles de su regulación e integración con otras vías metabólicas.
Secuencia de reacciones
Resumen de reacciones
Glucosa

HexoquinasaatpADP
Glucosa 6-fosfato

Glucosa-6-fosfatoisomerasa
Fructosa 6-fosfato

Fosfofructoquinasa-1atpADP
Fructosa 1,6-bifosfato

Fructosa-bifosfatoaldolasa
Fosfato de dihidroxiacetona

++
Gliceraldehído 3-fosfato

triosafosfatoisomerasa
2 × gliceraldehído 3-fosfato
2 ×
Gliceraldehído-3-fosfatodeshidrogenasaNAD +
PiNADH+H
NAD +
PiNADH+H
2 × 1,3-bisfosfoglicerato
2 × 
Fosfoglicerato quinasaADPatp
ADPatp
2 × 3-fosfoglicerato
2 × 
Fosfoglicerato mutasa
2 × 2-fosfoglicerato
2 × 
Fosfopiruvatohidratasa (enolasa)H2O
_ _
H2O
_ _
2 × fosfoenolpiruvato
2 × 
piruvato quinasaADPatp
2 × piruvato
2 × ![]()
Frase preparatoria
Los primeros cinco pasos de la glucólisis se consideran la fase preparatoria (o de inversión), ya que consumen energía para convertir la glucosa en dos azúcares fosfato de tres carbonos (G3P).
d -Glucosa (Glc)Hexoquinasa glucoquinasa (HK)una transferasaα - d -glucosa-6-fosfato (G6P) 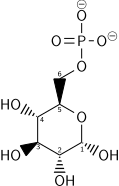 atpH +ADP atpH +ADP |
El primer paso es la fosforilación de la glucosa por una familia de enzimas llamadas hexocinasas para formar glucosa 6-fosfato (G6P). Esta reacción consume ATP, pero actúa para mantener baja la concentración de glucosa, promoviendo el transporte continuo de glucosa al interior de la célula a través de los transportadores de la membrana plasmática. Además, bloquea la fuga de glucosa: la célula carece de transportadores para G6P y se impide la libre difusión fuera de la célula debido a la naturaleza cargada de G6P. La glucosa se puede formar alternativamente a partir de la fosforólisis o hidrólisis de almidón o glucógeno intracelular.
En los animales, también se usa en el hígado una isoenzima de hexocinasa llamada glucocinasa, que tiene una afinidad mucho menor por la glucosa (K m en la vecindad de la glucemia normal) y difiere en sus propiedades reguladoras. La diferente afinidad por el sustrato y la regulación alternativa de esta enzima son un reflejo del papel del hígado en el mantenimiento de los niveles de azúcar en la sangre.
Cofactores: Mg
α - d -Glucosa 6-fosfato (G6P)Fosfoglucoisomerasa (PGI)una isomerasaβ - d -fructosa 6-fosfato (F6P)  |
Luego, la glucosa fosfato isomerasa transforma la G6P en fructosa 6-fosfato (F6P). La fructosa también puede entrar en la vía glucolítica por fosforilación en este punto.
El cambio de estructura es una isomerización, en la que el G6P se ha convertido en F6P. La reacción requiere una enzima, la fosfoglucosa isomerasa, para continuar. Esta reacción es libremente reversible en condiciones celulares normales. Sin embargo, a menudo se impulsa debido a una baja concentración de F6P, que se consume constantemente durante el siguiente paso de la glucólisis. En condiciones de alta concentración de F6P, esta reacción se desarrolla fácilmente a la inversa. Este fenómeno se puede explicar a través del Principio de Le Chatelier. La isomerización a un cetoazúcar es necesaria para la estabilización del carbanión en el cuarto paso de reacción (abajo).
β - d -fructosa 6-fosfato (F6P)Fosfofructoquinasa (PFK-1)una transferasaβ - d -fructosa 1,6-bisfosfato (F1,6BP) 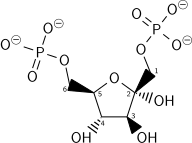 atpH +ADP atpH +ADP |
El gasto energético de otro ATP en este paso se justifica de 2 formas: El proceso glicolítico (hasta este paso) se vuelve irreversible, y la energía aportada desestabiliza la molécula. Debido a que la reacción catalizada por la fosfofructoquinasa 1 (PFK-1) está acoplada a la hidrólisis de ATP (un paso energéticamente favorable), es, en esencia, irreversible y se debe utilizar una vía diferente para realizar la conversión inversa durante la gluconeogénesis. Esto hace que la reacción sea un punto regulatorio clave (ver más abajo). Este es también el paso limitante de la velocidad.
Además, el segundo evento de fosforilación es necesario para permitir la formación de dos grupos cargados (en lugar de uno solo) en el paso posterior de la glucólisis, asegurando la prevención de la difusión libre de sustratos fuera de la célula.
La misma reacción también puede ser catalizada por la fosfofructoquinasa dependiente de pirofosfato (PFP o PPi-PFK), que se encuentra en la mayoría de las plantas, algunas bacterias, arqueas y protistas, pero no en los animales. Esta enzima utiliza pirofosfato (PPi) como donante de fosfato en lugar de ATP. Es una reacción reversible, aumentando la flexibilidad del metabolismo glucolítico. Se ha identificado una variante de enzima PFK dependiente de ADP más rara en especies de arqueas.
Cofactores: Mg
β - d -fructosa 1,6-bisfosfato (F1,6BP)Fructosa-bifosfato aldolasa (ALDO)una liasad -gliceraldehído 3-fosfato (GADP)Fosfato de dihidroxiacetona (DHAP) + + |
La desestabilización de la molécula en la reacción anterior permite que la aldolasa divida el anillo de hexosa en dos azúcares triosa: dihidroxiacetona fosfato (una cetosa) y gliceraldehído 3-fosfato (una aldosa). Hay dos clases de aldolasas: aldolasas de clase I, presentes en animales y plantas, y aldolasas de clase II, presentes en hongos y bacterias; las dos clases utilizan diferentes mecanismos para dividir el anillo de cetosa.
Los electrones deslocalizados en la escisión del enlace carbono-carbono se asocian con el grupo alcohol. El carbanión resultante está estabilizado por la estructura del propio carbanión a través de la distribución de carga de resonancia y por la presencia de un grupo prostético de iones cargados.
| Fosfato de dihidroxiacetona (DHAP)Triosafosfato isomerasa (TPI)una isomerasad -gliceraldehído 3-fosfato (GADP) |
La triosafosfato isomerasa interconvierte rápidamente el fosfato de dihidroxiacetona con gliceraldehído 3-fosfato (GADP) que continúa en la glucólisis. Esto es ventajoso, ya que dirige el fosfato de dihidroxiacetona por la misma vía que el 3-fosfato de gliceraldehído, lo que simplifica la regulación.
Fase de liquidación
La segunda mitad de la glucólisis se conoce como la fase de pago, caracterizada por una ganancia neta de moléculas ATP y NADH ricas en energía. Dado que la glucosa conduce a dos azúcares triosas en la fase preparatoria, cada reacción en la fase de pago ocurre dos veces por molécula de glucosa. Esto produce 2 moléculas de NADH y 4 moléculas de ATP, lo que lleva a una ganancia neta de 2 moléculas de NADH y 2 moléculas de ATP de la vía glucolítica por glucosa.
Gliceraldehído 3-fosfato (GADP)Gliceraldehído fosfato deshidrogenasa (GAPDH)una oxidorreductasad -1,3-bisfosfoglicerato (1,3BPG)  NAD + Pi _NADH + H NAD + Pi _NADH + H |
Los grupos aldehído de los azúcares de triosa se oxidan y se les añade fosfato inorgánico, formando 1,3-bisfosfoglicerato.
El hidrógeno se usa para reducir dos moléculas de NAD, un portador de hidrógeno, para dar NADH + H para cada triosa.
El equilibrio del átomo de hidrógeno y el equilibrio de carga se mantienen porque el grupo fosfato (P i) en realidad existe en forma de anión fosfato de hidrógeno (HPO 4), que se disocia para aportar el ion H adicional y da una carga neta de -3 en ambos lados
Aquí, el arsenato (AsO 4), un anión similar al fosfato inorgánico, puede reemplazar al fosfato como sustrato para formar 1-arseno-3-fosfoglicerato. Este, sin embargo, es inestable y se hidroliza fácilmente para formar 3-fosfoglicerato, el intermediario en el siguiente paso de la ruta. Como consecuencia de pasar por alto este paso, la molécula de ATP generada a partir del bisfosfoglicerato 1-3 en la siguiente reacción no se producirá, aunque la reacción prosiga. Como resultado, el arseniato es un desacoplador de la glucólisis.
1,3-bisfosfoglicerato (1,3BPG)Fosfoglicerato quinasa (PGK)una transferasa3-fosfoglicerato (3PG) 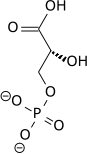 ADPatp ADPatp |
Este paso es la transferencia enzimática de un grupo fosfato del 1,3-bisfosfoglicerato al ADP por la fosfoglicerato quinasa, formando ATP y 3-fosfoglicerato. En este paso, la glucólisis ha alcanzado el punto de equilibrio: se consumieron 2 moléculas de ATP y ahora se han sintetizado 2 nuevas moléculas. Este paso, uno de los dos pasos de fosforilación a nivel de sustrato, requiere ADP; así, cuando la célula tiene mucho ATP (y poco ADP), esta reacción no ocurre. Debido a que el ATP se descompone con relativa rapidez cuando no se metaboliza, este es un punto regulador importante en la vía glucolítica.
ADP en realidad existe como ADPMg y ATP como ATPMg, equilibrando las cargas en −5 en ambos lados.
Cofactores: Mg
3-fosfoglicerato (3PG)Fosfoglicerato mutasa (PGM)una mutasa2-fosfoglicerato (2PG)  |
La fosfoglicerato mutasa isomeriza el 3-fosfoglicerato en 2-fosfoglicerato.
2-fosfoglicerato (2PG)Enolasa (ENO)una liasaFosfoenolpiruvato (PEP)  H2O _ _ H2O _ _ |
La enolasa luego convierte el 2-fosfoglicerato en fosfoenolpiruvato. Esta reacción es una reacción de eliminación que implica un mecanismo E1cB.
Cofactores: 2 Mg, un ion "conformacional" para coordinarse con el grupo carboxilato del sustrato, y un ion "catalítico" que participa en la deshidratación.
Fosfoenolpiruvato (PEP)Piruvato quinasa (PK)una transferasaPiruvato (Pyr)  ADP+Hatp ADP+Hatp |
Una fosforilación final a nivel de sustrato ahora forma una molécula de piruvato y una molécula de ATP por medio de la enzima piruvato quinasa. Esto sirve como un paso regulador adicional, similar al paso de la fosfoglicerato quinasa.
Cofactores: Mg
Lógica bioquímica
La existencia de más de un punto de regulación indica que los intermedios entre esos puntos entran y salen de la vía de la glucólisis por otros procesos. Por ejemplo, en el primer paso regulado, la hexocinasa convierte la glucosa en glucosa-6-fosfato. En lugar de continuar a través de la vía de la glucólisis, este intermediario puede convertirse en moléculas de almacenamiento de glucosa, como glucógeno o almidón. La reacción inversa, la descomposición, por ejemplo, del glucógeno, produce principalmente glucosa-6-fosfato; se forma muy poca glucosa libre en la reacción. La glucosa-6-fosfato así producida puede entrar en la glucólisis después del primer punto de control.
En el segundo paso regulado (el tercer paso de la glucólisis), la fosfofructocinasa convierte la fructosa-6-fosfato en fructosa-1,6-bifosfato, que luego se convierte en gliceraldehído-3-fosfato y dihidroxiacetona fosfato. El fosfato de dihidroxiacetona se puede eliminar de la glucólisis mediante la conversión en glicerol-3-fosfato, que se puede usar para formar triglicéridos. Por el contrario, los triglicéridos se pueden descomponer en ácidos grasos y glicerol; este último, a su vez, puede convertirse en fosfato de dihidroxiacetona, que puede entrar en la glucólisis después del segundo punto de control.
Cambios de energía libre
| Compuesto | Concentración / mM |
|---|---|
| Glucosa | 5.0 |
| Glucosa-6-fosfato | 0.083 |
| Fructosa-6-fosfato | 0.014 |
| Fructosa-1,6-bifosfato | 0.031 |
| Fosfato de dihidroxiacetona | 0.14 |
| Gliceraldehído-3-fosfato | 0.019 |
| 1,3-bisfosfoglicerato | 0.001 |
| 2,3-bisfosfoglicerato | 4.0 |
| 3-fosfoglicerato | 0.12 |
| 2-fosfoglicerato | 0.03 |
| Fosfoenolpiruvato | 0.023 |
| piruvato | 0.051 |
| atp | 1.85 |
| ADP | 0.14 |
| pi _ | 1.0 |
El cambio en la energía libre, Δ G, para cada paso en la vía de la glucólisis se puede calcular usando Δ G = Δ G °' + RT ln Q, donde Q es el cociente de reacción. Esto requiere conocer las concentraciones de los metabolitos. Todos estos valores están disponibles para eritrocitos, a excepción de las concentraciones de NAD y NADH. La proporción de NAD a NADH en el citoplasma es de aproximadamente 1000, lo que hace que la oxidación del gliceraldehído-3-fosfato (paso 6) sea más favorable.
Utilizando las concentraciones medidas de cada paso y los cambios de energía libre estándar, se puede calcular el cambio de energía libre real. (Descuidar esto es muy común: el delta G de la hidrólisis de ATP en las células no es el cambio de energía libre estándar de la hidrólisis de ATP citado en los libros de texto).
| Paso | Reacción | Δ G °' / (kJ/mol) | ΔG / (kJ /mol) |
|---|---|---|---|
| 1 | Glucosa + ATP → Glucosa-6-fosfato + ADP + H | −16,7 | −34 |
| 2 | Glucosa-6-fosfato → Fructosa-6-fosfato | 1.67 | −2,9 |
| 3 | Fructosa-6-fosfato + ATP → Fructosa-1,6-bisfosfato + ADP + H | −14,2 | −19 |
| 4 | Fructosa-1,6-bisfosfato → Dihidroxiacetona fosfato + Gliceraldehído-3-fosfato | 23,9 | −0,23 |
| 5 | Fosfato de dihidroxiacetona → Gliceraldehído-3-fosfato | 7.56 | 2.4 |
| 6 | Gliceraldehído-3-fosfato + P i + NAD → 1,3-bisfosfoglicerato + NADH + H | 6.30 | −1,29 |
| 7 | 1,3-Bisfosfoglicerato + ADP → 3-Fosfoglicerato + ATP | −18,9 | 0.09 |
| 8 | 3-fosfoglicerato → 2-fosfoglicerato | 4.4 | 0.83 |
| 9 | 2-fosfoglicerato → fosfoenolpiruvato + H 2 O | 1.8 | 1.1 |
| 10 | Fosfoenolpiruvato + ADP + H → Piruvato + ATP | −31,7 | −23,0 |
A partir de la medición de las concentraciones fisiológicas de metabolitos en un eritrocito, parece que alrededor de siete de los pasos de la glucólisis están en equilibrio para ese tipo de célula. Tres de los pasos, los que tienen grandes cambios negativos de energía libre, no están en equilibrio y se denominan irreversibles; dichos pasos a menudo están sujetos a regulación.
El paso 5 en la figura se muestra detrás de los otros pasos, porque ese paso es una reacción secundaria que puede disminuir o aumentar la concentración del intermediario gliceraldehído-3-fosfato. Ese compuesto se convierte en fosfato de dihidroxiacetona por la enzima triosa fosfato isomerasa, que es una enzima catalíticamente perfecta; su velocidad es tan rápida que se puede suponer que la reacción está en equilibrio. El hecho de que ΔG no sea cero indica que las concentraciones reales en el eritrocito no se conocen con precisión.
Regulación
Las enzimas son los componentes principales que impulsan la vía metabólica y, por lo tanto, la exploración de los mecanismos reguladores de estas enzimas nos dará una idea de los procesos reguladores que afectan a la glucólisis. Hay un total de 9 pasos principales en la glucólisis que es impulsada por 14 enzimas diferentes. Las enzimas pueden modificarse o verse afectadas mediante 5 procesos reguladores principales, incluida la modificación postraduccional (PTM) y la localización.
Mecanismos biológicos por los que se regulan las enzimas
1. Expresión génica2. Alosteria3. Interacción proteína-proteína (PPI)4. Modificación postraduccional (PTM)5. Localización
Regulación por insulina en animales
En los animales, la regulación de los niveles de glucosa en sangre por parte del páncreas junto con el hígado es una parte vital de la homeostasis. Las células beta en los islotes pancreáticos son sensibles a la concentración de glucosa en sangre. Un aumento en la concentración de glucosa en sangre hace que liberen insulina en la sangre, lo que tiene un efecto particularmente en el hígado, pero también en las células grasas y musculares, lo que hace que estos tejidos eliminen la glucosa de la sangre. Cuando el azúcar en la sangre cae, las células beta pancreáticas dejan de producir insulina, pero, en cambio, estimulan a las células alfa pancreáticas vecinas para que liberen glucagón en la sangre.Esto, a su vez, hace que el hígado libere glucosa a la sangre al descomponer el glucógeno almacenado y por medio de la gluconeogénesis. Si la caída en el nivel de glucosa en sangre es particularmente rápida o severa, otros sensores de glucosa provocan la liberación de epinefrina de las glándulas suprarrenales a la sangre. Tiene la misma acción que el glucagón sobre el metabolismo de la glucosa, pero su efecto es más pronunciado. En el hígado, el glucagón y la epinefrina provocan la fosforilación de las enzimas clave limitantes de la velocidad de la glucólisis, la síntesis de ácidos grasos, la síntesis de colesterol, la gluconeogénesis y la glucogenólisis. La insulina tiene el efecto opuesto sobre estas enzimas.La fosforilación y desfosforilación de estas enzimas (en última instancia, en respuesta al nivel de glucosa en la sangre) es la forma dominante mediante la cual se controlan estas vías en las células hepáticas, grasas y musculares. Así, la fosforilación de la fosfofructocinasa inhibe la glucólisis, mientras que su desfosforilación por acción de la insulina estimula la glucólisis.
Regulación de las enzimas limitantes de la velocidad
Las tres enzimas reguladoras son hexoquinasa (o glucoquinasa en el hígado), fosfofructoquinasa y piruvato quinasa. El flujo a través de la vía glucolítica se ajusta en respuesta a las condiciones tanto dentro como fuera de la célula. Los factores internos que regulan la glucólisis lo hacen principalmente para proporcionar ATP en cantidades adecuadas para las necesidades de la célula. Los factores externos actúan principalmente sobre el hígado, el tejido adiposo y los músculos, que pueden eliminar grandes cantidades de glucosa de la sangre después de las comidas (previniendo así la hiperglucemia al almacenar el exceso de glucosa como grasa o glucógeno, según el tipo de tejido). El hígado también es capaz de liberar glucosa a la sangre entre comidas, durante el ayuno y el ejercicio, evitando así la hipoglucemia mediante la glucogenólisis y la gluconeogénesis.
Además, la hexoquinasa y la glucoquinasa actúan independientemente de los efectos hormonales como controles en los puntos de entrada de la glucosa a las células de diferentes tejidos. La hexoquinasa responde al nivel de glucosa-6-fosfato (G6P) en la célula o, en el caso de la glucoquinasa, al nivel de azúcar en la sangre para impartir controles completamente intracelulares de la vía glucolítica en diferentes tejidos (ver más abajo).
Cuando la glucosa ha sido convertida en G6P por hexocinasa o glucocinasa, puede convertirse en glucosa-1-fosfato (G1P) para convertirla en glucógeno, o alternativamente se convierte mediante glucólisis en piruvato, que ingresa a la mitocondria donde se convierte en acetil-CoA y luego en citrato. El exceso de citrato se exporta de la mitocondria al citosol, donde la ATP citrato liasa regenera acetil-CoA y oxaloacetato (OAA). El acetil-CoA se usa luego para la síntesis de ácidos grasos y la síntesis de colesterol, dos formas importantes de utilizar el exceso de glucosa cuando su concentración es alta en la sangre. Las enzimas limitantes de la velocidad que catalizan estas reacciones realizan estas funciones cuando han sido desfosforiladas mediante la acción de la insulina sobre las células hepáticas. Entre comidas, durante el ayuno, ejercicio o hipoglucemia, el glucagón y la epinefrina se liberan en la sangre. Esto hace que el glucógeno hepático se convierta de nuevo en G6P y luego se convierta en glucosa mediante la enzima específica del hígado glucosa 6-fosfatasa y se libere en la sangre. El glucagón y la epinefrina también estimulan la gluconeogénesis, que convierte los sustratos que no son carbohidratos en G6P, que se une a la G6P derivada del glucógeno, o la sustituye cuando se agotan las reservas de glucógeno del hígado. Esto es fundamental para la función cerebral, ya que el cerebro utiliza la glucosa como fuente de energía en la mayoría de las condiciones. que convierte los sustratos que no son carbohidratos en G6P, que se une a la G6P derivada del glucógeno, o la sustituye cuando las reservas de glucógeno hepático se han agotado. Esto es fundamental para la función cerebral, ya que el cerebro utiliza la glucosa como fuente de energía en la mayoría de las condiciones. que convierte los sustratos que no son carbohidratos en G6P, que se une a la G6P derivada del glucógeno, o la sustituye cuando las reservas de glucógeno hepático se han agotado. Esto es fundamental para la función cerebral, ya que el cerebro utiliza la glucosa como fuente de energía en la mayoría de las condiciones.La fosforilación simultánea de, en particular, la fosfofructoquinasa, pero también, en cierta medida, la piruvato quinasa, impide que la glucólisis se produzca al mismo tiempo que la gluconeogénesis y la glucogenólisis.
Hexocinasa y glucocinasa
Todas las células contienen la enzima hexocinasa, que cataliza la conversión de la glucosa que ha entrado en la célula en glucosa-6-fosfato (G6P). Dado que la membrana celular es impermeable a la G6P, la hexoquinasa actúa esencialmente para transportar glucosa a las células de las que ya no puede escapar. La hexoquinasa es inhibida por altos niveles de G6P en la célula. Por lo tanto, la tasa de entrada de glucosa en las células depende parcialmente de la rapidez con la que se puede eliminar la G6P mediante la glucólisis y la síntesis de glucógeno (en las células que almacenan glucógeno, es decir, el hígado y los músculos).
La glucoquinasa, a diferencia de la hexoquinasa, no es inhibida por G6P. Ocurre en las células hepáticas y solo fosforila la glucosa que ingresa a la célula para formar glucosa-6-fosfato (G6P), cuando la glucosa en la sangre es abundante. Siendo este el primer paso en la ruta glucolítica en el hígado, imparte una capa adicional de control de la ruta glucolítica en este órgano.
Fosfofructoquinasa
La fosfofructocinasa es un punto de control importante en la vía glucolítica, ya que es uno de los pasos irreversibles y tiene efectores alostéricos clave, AMP y fructosa 2,6-bisfosfato (F2,6BP).
La fructosa 2,6-bisfosfato (F2,6BP) es un activador muy potente de la fosfofructoquinasa (PFK-1) que se sintetiza cuando la F6P es fosforilada por una segunda fosfofructoquinasa (PFK2). En el hígado, cuando el azúcar en la sangre es bajo y el glucagón eleva el AMPc, la proteína quinasa A fosforila la PFK2. La fosforilación inactiva la PFK2 y otro dominio de esta proteína se activa como fructosa bisfosfatasa-2, que convierte F2,6BP nuevamente en F6P. Tanto el glucagón como la epinefrina provocan altos niveles de AMPc en el hígado. El resultado de niveles más bajos de fructosa-2,6-bifosfato hepático es una disminución de la actividad de la fosfofructocinasa y un aumento de la actividad de la fructosa 1,6-bisfosfatasa, de modo que se favorece la gluconeogénesis (en esencia, "glucólisis a la inversa"). Esto es consistente con el papel del hígado en tales situaciones,
ATP compite con AMP por el sitio efector alostérico en la enzima PFK. Las concentraciones de ATP en las células son mucho más altas que las de AMP, generalmente 100 veces más altas, pero la concentración de ATP no cambia más de un 10 % en condiciones fisiológicas, mientras que una caída del 10 % en ATP resulta en un aumento de 6 veces en AMPERIO. Por lo tanto, la relevancia del ATP como efector alostérico es cuestionable. Un aumento en AMP es consecuencia de una disminución en la carga de energía en la célula.
El citrato inhibe la fosfofructocinasa cuando se prueba in vitro al aumentar el efecto inhibidor del ATP. Sin embargo, es dudoso que este sea un efecto significativo in vivo, porque el citrato en el citosol se utiliza principalmente para la conversión a acetil-CoA para la síntesis de ácidos grasos y colesterol.
TIGAR, una enzima inducida por p53, es responsable de la regulación de la fosfofructoquinasa y actúa para proteger contra el estrés oxidativo. TIGAR es una enzima única con doble función que regula F2,6BP. Puede comportarse como una fosfatasa (fructosa-2,6-bisfosfatasa) que escinde el fosfato en el carbono-2 produciendo F6P. También puede comportarse como una quinasa (PFK2) agregando un fosfato al carbono-2 de F6P que produce F2,6BP. En humanos, la proteína TIGAR está codificada por el gen C12orf5. La enzima TIGAR obstaculizará el avance de la glucólisis al crear una acumulación de fructosa-6-fosfato (F6P) que se isomeriza en glucosa-6-fosfato (G6P). La acumulación de G6P desviará los carbonos hacia la ruta de las pentosas fosfato.
Piruvato quinasa
La enzima piruvato quinasa cataliza el último paso de la glucólisis, en el que se forman piruvato y ATP. La piruvato quinasa cataliza la transferencia de un grupo fosfato del fosfoenolpiruvato (PEP) al ADP, produciendo una molécula de piruvato y una molécula de ATP.
La piruvato cinasa hepática está regulada indirectamente por la epinefrina y el glucagón, a través de la proteína cinasa A. Esta proteína cinasa fosforila la piruvato cinasa hepática para desactivarla. La piruvato quinasa muscular no es inhibida por la activación de la proteína quinasa A por parte de la epinefrina. El glucagón señala el ayuno (no hay glucosa disponible). Por lo tanto, la glucólisis se inhibe en el hígado pero no se ve afectada en el músculo en ayunas. Un aumento en el azúcar en la sangre conduce a la secreción de insulina, que activa la fosfoproteína fosfatasa I, lo que lleva a la desfosforilación y activación de la piruvato quinasa. Estos controles evitan que la piruvato quinasa esté activa al mismo tiempo que las enzimas que catalizan la reacción inversa (piruvato carboxilasa y fosfoenolpiruvato carboxicinasa), impidiendo un ciclo fútil.
Procesos posteriores a la glucólisis
El proceso general de la glucólisis es:Glucosa + 2 NAD + 2 ADP + 2 P i → 2 piruvato + 2 NADH + 2 H + 2 ATP
Si la glucólisis continuara indefinidamente, todo el NAD se consumiría y la glucólisis se detendría. Para permitir que continúe la glucólisis, los organismos deben poder oxidar el NADH de nuevo a NAD. Cómo se realiza esto depende de qué aceptor de electrones externo esté disponible.
Regeneración anóxica de NAD
Un método para hacer esto es simplemente hacer que el piruvato haga la oxidación; en este proceso, el piruvato se convierte en lactato (la base conjugada del ácido láctico) en un proceso llamado fermentación del ácido láctico:Piruvato + NADH + H → lactato + NAD
Este proceso ocurre en las bacterias involucradas en la elaboración del yogur (el ácido láctico hace que la leche cuaje). Este proceso también ocurre en animales bajo condiciones hipóxicas (o parcialmente anaeróbicas), que se encuentran, por ejemplo, en músculos sobrecargados que carecen de oxígeno. En muchos tejidos, este es un último recurso celular para obtener energía; la mayoría de los tejidos animales no pueden tolerar las condiciones anaeróbicas durante un período de tiempo prolongado.
Algunos organismos, como la levadura, convierten el NADH nuevamente en NAD en un proceso llamado fermentación de etanol. En este proceso, el piruvato se convierte primero en acetaldehído y dióxido de carbono, y luego en etanol.
La fermentación de ácido láctico y la fermentación de etanol pueden ocurrir en ausencia de oxígeno. Esta fermentación anaeróbica permite que muchos organismos unicelulares utilicen la glucólisis como su única fuente de energía.
La regeneración anóxica de NAD es solo un medio efectivo de producción de energía durante el ejercicio intenso y corto en vertebrados, por un período que varía de 10 segundos a 2 minutos durante un esfuerzo máximo en humanos. (A intensidades de ejercicio más bajas, puede mantener la actividad muscular en animales buceadores, como focas, ballenas y otros vertebrados acuáticos, durante períodos de tiempo mucho más largos). Bajo estas condiciones NADse repone con NADH que dona sus electrones al piruvato para formar lactato. Esto produce 2 moléculas de ATP por molécula de glucosa, o alrededor del 5% del potencial energético de la glucosa (38 moléculas de ATP en bacterias). Pero la velocidad a la que se produce ATP de esta manera es unas 100 veces mayor que la de la fosforilación oxidativa. El pH en el citoplasma cae rápidamente cuando los iones de hidrógeno se acumulan en el músculo, lo que eventualmente inhibe las enzimas involucradas en la glucólisis.
La sensación de ardor en los músculos durante el ejercicio intenso se puede atribuir a la liberación de iones de hidrógeno durante el cambio a la fermentación de la glucosa desde la oxidación de la glucosa hasta el dióxido de carbono y el agua, cuando el metabolismo aeróbico ya no puede seguir el ritmo de las demandas de energía de los músculos. Estos iones de hidrógeno forman parte del ácido láctico. El cuerpo recurre a este método menos eficiente pero más rápido de producir ATP en condiciones de bajo nivel de oxígeno. Se cree que este fue el principal medio de producción de energía en organismos anteriores antes de que el oxígeno alcanzara altas concentraciones en la atmósfera hace entre 2000 y 2500 millones de años y, por lo tanto, representaría una forma más antigua de producción de energía que la reposición aeróbica de NAD en las células..
El hígado de los mamíferos se deshace de este exceso de lactato transformándolo nuevamente en piruvato en condiciones aeróbicas; ver ciclo de Cori.
La fermentación de piruvato a lactato a veces también se denomina "glucólisis anaeróbica", sin embargo, la glucólisis termina con la producción de piruvato independientemente de la presencia o ausencia de oxígeno.
En los dos ejemplos de fermentación anteriores, el NADH se oxida mediante la transferencia de dos electrones al piruvato. Sin embargo, las bacterias anaerobias utilizan una amplia variedad de compuestos como aceptores terminales de electrones en la respiración celular: compuestos nitrogenados, como nitratos y nitritos; compuestos de azufre, tales como sulfatos, sulfitos, dióxido de azufre y azufre elemental; dióxido de carbono; compuestos de hierro; compuestos de manganeso; compuestos de cobalto; y compuestos de uranio.
Regeneración aeróbica de NAD y eliminación de piruvato.
En los organismos aeróbicos, se ha desarrollado un mecanismo complejo para utilizar el oxígeno del aire como aceptor final de electrones.
- En primer lugar, el NADH + H generado por la glucólisis tiene que ser transferido a la mitocondria para ser oxidado y así regenerar el NAD necesario para que continúe la glucólisis. Sin embargo, la membrana mitocondrial interna es impermeable a NADH y NAD. Por lo tanto, se utilizan dos "lanzaderas" para transportar los electrones del NADH a través de la membrana mitocondrial. Son la lanzadera de malato-aspartato y la lanzadera de fosfato de glicerol. En el primero, los electrones del NADH se transfieren al oxalacetato citosólico para formar malato. El malato luego atraviesa la membrana mitocondrial interna hacia la matriz mitocondrial, donde es reoxidado por NAD.formando oxaloacetato intramitocondrial y NADH. Luego, el oxaloacetato se recicla al citosol a través de su conversión en aspartato, que se transporta fácilmente fuera de la mitocondria. En la lanzadera de glicerol fosfato, los electrones del NADH citosólico se transfieren a la dihidroxiacetona para formar glicerol-3-fosfato que atraviesa fácilmente la membrana mitocondrial externa. Luego, el glicerol-3-fosfato se reoxida a dihidroxiacetona, donando sus electrones a FAD en lugar de NAD. Esta reacción tiene lugar en la membrana mitocondrial interna, lo que permite que FADH 2 done sus electrones directamente a la coenzima Q (ubiquinona), que forma parte de la cadena de transporte de electrones que finalmente transfiere electrones al oxígeno molecular (O 2), con la formación de agua y la liberación de energía eventualmente capturada en forma de ATP.
- El producto final glucolítico, el piruvato (más NAD), se convierte en acetil-CoA, CO 2 y NADH + H dentro de las mitocondrias en un proceso llamado descarboxilación del piruvato.
- El acetil-CoA resultante ingresa al ciclo del ácido cítrico (o ciclo de Krebs), donde el grupo acetilo del acetil-CoA se convierte en dióxido de carbono mediante dos reacciones de descarboxilación con la formación de aún más NADH + H intramitocondrial.
- El NADH + H intramitocondrial se oxida a NAD por la cadena de transporte de electrones, utilizando oxígeno como aceptor final de electrones para formar agua. La energía liberada durante este proceso se utiliza para crear un gradiente de iones de hidrógeno (o protones) a través de la membrana interna de la mitocondria.
- Finalmente, el gradiente de protones se usa para producir alrededor de 2.5 ATP por cada NADH + H oxidado en un proceso llamado fosforilación oxidativa.
Conversión de carbohidratos en ácidos grasos y colesterol
El piruvato producido por la glucólisis es un intermediario importante en la conversión de carbohidratos en ácidos grasos y colesterol. Esto ocurre a través de la conversión de piruvato en acetil-CoA en la mitocondria. Sin embargo, este acetil CoA necesita ser transportado al citosol donde ocurre la síntesis de ácidos grasos y colesterol. Esto no puede ocurrir directamente. Para obtener acetil-CoA citosólico, el citrato (producido por la condensación de acetil CoA con oxaloacetato) se elimina del ciclo del ácido cítrico y se transporta a través de la membrana mitocondrial interna hacia el citosol.Allí es escindido por ATP citrato liasa en acetil-CoA y oxaloacetato. El oxaloacetato se devuelve a la mitocondria como malato (y luego vuelve al oxaloacetato para transferir más acetil-CoA fuera de la mitocondria). La acetil-CoA citosólica puede ser carboxilada por la acetil-CoA carboxilasa en malonil CoA, el primer paso comprometido en la síntesis de ácidos grasos, o puede combinarse con acetoacetil-CoA para formar 3-hidroxi-3-metilglutaril-CoA (HMG -CoA), que es el paso limitante de la velocidad que controla la síntesis de colesterol. El colesterol se puede usar tal cual, como componente estructural de las membranas celulares, o se puede usar para sintetizar las hormonas esteroides, las sales biliares y la vitamina D.
Conversión de piruvato en oxaloacetato para el ciclo del ácido cítrico
Las moléculas de piruvato producidas por la glucólisis se transportan activamente a través de la membrana mitocondrial interna y hacia la matriz, donde pueden oxidarse y combinarse con la coenzima A para formar CO 2, acetil-CoA y NADH, o pueden carboxilarse (mediante la piruvato carboxilasa).) para formar oxalacetato. Esta última reacción "llena" la cantidad de oxaloacetato en el ciclo del ácido cítrico, y por tanto es una reacción anaplerótica (del griego que significa "llenar"), aumentando la capacidad del ciclo para metabolizar acetil-CoA cuando el tejido necesita energía (ej., en el corazón y el músculo esquelético) aumentan repentinamente por la actividad. En el ciclo del ácido cítrico, todos los intermedios (p. ej., citrato, isocitrato, alfa-cetoglutarato, succinato, fumarato, malato y oxaloacetato) se regeneran durante cada turno del ciclo. Por lo tanto, agregar más de cualquiera de estos intermediarios a la mitocondria significa que esa cantidad adicional se retiene dentro del ciclo, aumentando todos los demás intermediarios a medida que uno se convierte en el otro. Por lo tanto, la adición de oxaloacetato aumenta considerablemente las cantidades de todos los intermediarios del ácido cítrico, lo que aumenta la capacidad del ciclo para metabolizar la acetil CoA, convirtiendo su componente acetato en CO 2 y agua, con la liberación de energía suficiente para formar 11 moléculas de ATP y 1 de GTP. por cada molécula adicional de acetil CoA que se combina con oxaloacetato en el ciclo.
Para eliminar catapleróticamente el oxaloacetato del ciclo cítrico, el malato se puede transportar desde la mitocondria al citoplasma, lo que disminuye la cantidad de oxaloacetato que se puede regenerar. Además, los intermedios de ácido cítrico se utilizan constantemente para formar una variedad de sustancias como las purinas, pirimidinas y porfirinas.
Intermedios para otras vías
Este artículo se concentra en el papel catabólico de la glucólisis con respecto a la conversión de energía química potencial en energía química utilizable durante la oxidación de glucosa a piruvato. Muchos de los metabolitos en la ruta glucolítica también son utilizados por las rutas anabólicas y, como consecuencia, el flujo a través de la ruta es fundamental para mantener un suministro de esqueletos de carbono para la biosíntesis.
Las siguientes vías metabólicas dependen en gran medida de la glucólisis como fuente de metabolitos: y muchas más.
- La vía de las pentosas fosfato, que comienza con la deshidrogenación de glucosa-6-fosfato, el primer intermediario producido por la glucólisis, produce varios azúcares pentosos y NADPH para la síntesis de ácidos grasos y colesterol.
- La síntesis de glucógeno también comienza con glucosa-6-fosfato al comienzo de la vía glucolítica.
- El glicerol, para la formación de triglicéridos y fosfolípidos, se produce a partir del intermedio glucolítico gliceraldehído-3-fosfato.
- Varias vías post-glucolíticas:
- Síntesis de ácidos grasos
- Síntesis de colesterol
- El ciclo del ácido cítrico que a su vez conduce a:
- Síntesis de aminoácidos
- Síntesis de nucleótidos
- Síntesis de tetrapirrol
Aunque la gluconeogénesis y la glucólisis comparten muchos intermediarios, uno no es funcionalmente una rama o afluente del otro. Hay dos pasos reguladores en ambas vías que, cuando están activos en una vía, automáticamente están inactivos en la otra. Por lo tanto, los dos procesos no pueden estar activos simultáneamente. De hecho, si ambos conjuntos de reacciones fueran muy activos al mismo tiempo, el resultado neto sería la hidrólisis de cuatro enlaces fosfato de alta energía (dos ATP y dos GTP) por ciclo de reacción.
El NAD es el agente oxidante en la glucólisis, como lo es en la mayoría de las demás reacciones metabólicas generadoras de energía (p. ej., beta-oxidación de ácidos grasos y durante el ciclo del ácido cítrico). El NADH así producido se usa principalmente para transferir electrones a O 2 para producir agua o, cuando no hay O 2 disponible, para producir compuestos como lactato o etanol (ver Regeneración anóxica de NAD arriba). El NADH rara vez se usa para procesos sintéticos, con la notable excepción de la gluconeogénesis. Durante la síntesis de ácidos grasos y colesterol, el agente reductor es NADPH. Esta diferencia ejemplifica un principio general de que el NADPH se consume durante las reacciones biosintéticas, mientras que el NADH se genera en las reacciones que producen energía.La fuente del NADPH es doble. Cuando el malato se descarboxila oxidativamente por el piruvato de la "enzima málica unida a NADP", se forman CO 2 y NADPH. NADPH también se forma por la ruta de las pentosas fosfato que convierte la glucosa en ribosa, que puede usarse en la síntesis de nucleótidos y ácidos nucleicos, o puede catabolizarse a piruvato.
Glucólisis en la enfermedad
Diabetes
La captación celular de glucosa se produce en respuesta a las señales de insulina y, posteriormente, la glucosa se descompone a través de la glucólisis, lo que reduce los niveles de azúcar en la sangre. Sin embargo, los bajos niveles de insulina que se observan en la diabetes dan como resultado hiperglucemia, donde los niveles de glucosa en la sangre aumentan y las células no absorben la glucosa de manera adecuada. Los hepatocitos contribuyen aún más a esta hiperglucemia a través de la gluconeogénesis. La glucólisis en los hepatocitos controla la producción hepática de glucosa, y cuando el hígado produce un exceso de glucosa sin que el cuerpo la descomponga, se produce hiperglucemia.
Enfermedades genéticas
Las mutaciones glicolíticas son generalmente raras debido a la importancia de la ruta metabólica, esto significa que la mayoría de las mutaciones que ocurren dan como resultado una incapacidad para que la célula respire y, por lo tanto, causan la muerte de la célula en una etapa temprana. Sin embargo, se observan algunas mutaciones, siendo un ejemplo notable la deficiencia de piruvato quinasa, que conduce a la anemia hemolítica crónica.
Cáncer
Las células tumorales malignas realizan la glucólisis a un ritmo diez veces más rápido que sus contrapartes de tejido no canceroso. Durante su génesis, el apoyo capilar limitado a menudo da como resultado hipoxia (disminución del suministro de O2) dentro de las células tumorales. Por lo tanto, estas células dependen de procesos metabólicos anaeróbicos como la glucólisis para ATP (trifosfato de adenosina). Algunas células tumorales sobreexpresan enzimas glucolíticas específicas que dan como resultado tasas más altas de glucólisis. A menudo, estas enzimas son isoenzimas, de enzimas de glucólisis tradicionales, que varían en su susceptibilidad a la inhibición por retroalimentación tradicional. El aumento de la actividad glucolítica finalmente contrarresta los efectos de la hipoxia al generar suficiente ATP a partir de esta vía anaeróbica.Este fenómeno fue descrito por primera vez en 1930 por Otto Warburg y se conoce como efecto Warburg. La hipótesis de Warburg afirma que el cáncer es causado principalmente por una disfunción en el metabolismo mitocondrial, más que por el crecimiento descontrolado de las células. Se han propuesto varias teorías para explicar el efecto Warburg. Una de esas teorías sugiere que el aumento de la glucólisis es un proceso protector normal del cuerpo y que el cambio maligno podría ser causado principalmente por el metabolismo energético.
Esta alta tasa de glucólisis tiene importantes aplicaciones médicas, ya que la alta glucólisis aeróbica de los tumores malignos se utiliza clínicamente para diagnosticar y monitorear las respuestas al tratamiento de los cánceres mediante la captación de imágenes de 2-F-2-desoxiglucosa (FDG) (un sustrato de hexoquinasa modificada radioactiva) con positrones. tomografía por emisión (PET).
Hay investigaciones en curso para afectar el metabolismo mitocondrial y tratar el cáncer al reducir la glucólisis y, por lo tanto, matar de hambre a las células cancerosas de varias formas nuevas, incluida una dieta cetogénica.
Contenido relacionado
Baquelita
Experimento de Miller-Urey
Ave lira