
Flagelo
Un flagellum (pl. flagella) es un apéndice parecido a un cabello que sobresale de ciertos espermatozoides de plantas y animales, y de una amplia gama de microorganismos para proporcionar motilidad. Muchos protistas con flagelos se denominan flagelados.
Un microorganismo puede tener de uno a muchos flagelos. Una bacteria gramnegativa, Helicobacter pylori, por ejemplo, utiliza sus múltiples flagelos para impulsarse a través del revestimiento mucoso hasta llegar al epitelio del estómago, donde puede provocar el desarrollo de una úlcera gástrica. En algunas bacterias, el flagelo también puede funcionar como un orgánulo sensorial, siendo sensible a la humedad fuera de la célula.
En los tres dominios de bacterias, arqueas y eucariotas, el flagelo tiene una estructura, una composición proteica y un mecanismo de propulsión diferentes, pero comparte la misma función de proporcionar motilidad. La palabra latina flagellum significa "látigo" para describir su movimiento de natación similar a un látigo. El flagelo en archaea se llama archaellum para notar su diferencia con el flagelo bacteriano.
Los flagelos y cilios eucarióticos son idénticos en estructura pero tienen diferentes longitudes y funciones. Las fimbrias y pilosidades procarióticas son apéndices más pequeños y delgados, con diferentes funciones.
Tipos

Los tres tipos de flagelos son bacterianos, arqueales y eucariotas.
Los flagelos de los eucariotas tienen dineína y microtúbulos que se mueven con un mecanismo de flexión. Las bacterias y las arqueas no tienen dineína ni microtúbulos en sus flagelos y se mueven mediante un mecanismo giratorio.
Otras diferencias entre estos tres tipos son:
- La flagelación bacteriana son filamentos helicoidales, cada uno con un motor giratorio en su base que puede girar en sentido de reloj o en sentido contrario. Proporcionan dos de varios tipos de motilidad bacteriana.
- La flagella arqueal (archaella) es superficialmente similar a la flagella bacteriana, ya que también tiene un motor rotativo, pero son diferentes en muchos detalles y considerados no homólogos.
- La flagelo Eukaryotic —aquellas de células animales, vegetales y protistas— son proyecciones celulares complejas que se extienden hacia atrás y hacia adelante. La flagella Eukaryotic y los cilias motiles son idénticos en la estructura, pero tienen diferentes longitudes, formas de onda y funciones. Los cilias primarios son inmóviles, y tienen un axoneme estructuralmente diferente 9+0 en lugar del axoneme 9+2 encontrado en flagella y cilia fútil.
Bacteriana
(feminine)Estructura y composición
El flagelo bacteriano está formado por subunidades proteicas de flagelina. Su forma es un tubo hueco de 20 nanómetros de espesor. Es helicoidal y tiene una curva pronunciada justo fuera de la membrana exterior; este "gancho" permite que el eje de la hélice apunte directamente alejándose de la celda. Un eje corre entre el gancho y el cuerpo basal, pasando a través de anillos de proteína en la membrana de la célula que actúan como cojinetes. Los organismos grampositivos tienen dos de estos anillos corporales basales, uno en la capa de peptidoglicano y otro en la membrana plasmática. Los organismos gramnegativos tienen cuatro de estos anillos: el anillo L se asocia con los lipopolisacáridos, el anillo P se asocia con la capa de peptidoglucano, el anillo M está incrustado en la membrana plasmática y el anillo S está directamente unido a la membrana plasmática. El filamento termina con una proteína protectora.
El filamento flagelar es el tornillo helicoidal largo que impulsa a la bacteria cuando el motor la hace girar a través del gancho. En la mayoría de las bacterias que se han estudiado, incluidas las gramnegativas Escherichia coli, Salmonella typhimurium, Caulobacter crescentus y Vibrio alginolyticus, el filamento está formado por 11 protofilamentos aproximadamente paralelos al eje del filamento. Cada protofilamento es una serie de cadenas de proteínas en tándem. Sin embargo, Campylobacter jejuni tiene siete protofilamentos.
El cuerpo basal tiene varias características en común con algunos tipos de poros secretores, como el "tapón" en sus centros extendiéndose a través de la membrana plasmática. Las similitudes entre los flagelos bacterianos y las estructuras y proteínas del sistema secretor bacteriano proporcionan evidencia científica que respalda la teoría de que los flagelos bacterianos evolucionaron a partir del sistema de secreción tipo tres.
Motor
El flagelo bacteriano es impulsado por un motor rotatorio (complejo Mot) compuesto de proteína, ubicado en el punto de anclaje del flagelo en la membrana celular interna. El motor es impulsado por la fuerza motriz de protones, es decir, por el flujo de protones (iones de hidrógeno) a través de la membrana de la célula bacteriana debido a un gradiente de concentración establecido por el metabolismo de la célula (Vibrio especies tienen dos tipos de flagelos, laterales y polares, y algunos son impulsados por una bomba de iones de sodio en lugar de una bomba de protones). El rotor transporta protones a través de la membrana y gira en el proceso. El rotor por sí solo puede funcionar de 6000 a 17 000 rpm, pero con el filamento flagelar conectado, por lo general solo alcanza de 200 a 1000 rpm. La dirección de rotación puede ser cambiada por el interruptor del motor flagelar casi instantáneamente, causado por un ligero cambio en la posición de una proteína, FliG, en el rotor. El flagelo es muy eficiente energéticamente y utiliza muy poca energía. El mecanismo exacto para la generación de par aún no se conoce bien. Debido a que el motor flagelar no tiene un interruptor de encendido y apagado, la proteína epsE se usa como un embrague mecánico para desconectar el motor del rotor, deteniendo así el flagelo y permitiendo que la bacteria permanezca en un lugar.
La forma cilíndrica de los flagelos es adecuada para la locomoción de organismos microscópicos; estos organismos operan con un número de Reynolds bajo, donde la viscosidad del agua circundante es mucho más importante que su masa o inercia.
La velocidad de rotación de los flagelos varía en respuesta a la intensidad de la fuerza protón-motriz, lo que permite ciertas formas de control de la velocidad y también permite que algunos tipos de bacterias alcancen velocidades notables en proporción a su tamaño; algunos alcanzan aproximadamente 60 longitudes de celda por segundo. A tal velocidad, una bacteria tardaría unos 245 días en recorrer 1 km; aunque parezca lento, la perspectiva cambia cuando se introduce el concepto de escala. En comparación con las formas de vida macroscópicas, es realmente muy rápido cuando se expresa en términos de número de longitudes corporales por segundo. Un guepardo, por ejemplo, solo alcanza unas 25 longitudes corporales por segundo.
Mediante el uso de sus flagelos, las bacterias pueden moverse rápidamente hacia los atrayentes y alejarse de los repelentes, por medio de una caminata aleatoria sesgada, con 'carreras' y 'cae' provocada por la rotación de su flagelo en sentido antihorario y horario, respectivamente. Las dos direcciones de rotación no son idénticas (con respecto al movimiento del flagelo) y se seleccionan mediante un interruptor molecular.
Montaje
Durante el ensamblaje flagelar, los componentes del flagelo pasan a través de los núcleos huecos del cuerpo basal y el filamento naciente. Durante el ensamblaje, los componentes proteicos se agregan en la punta flagelar en lugar de en la base. In vitro, los filamentos flagelares se ensamblan espontáneamente en una solución que contiene flagelina purificada como única proteína.
Evolución
Al menos 10 componentes proteicos del flagelo bacteriano comparten proteínas homólogas con el sistema de secreción de tipo tres (T3SS) que se encuentra en muchas bacterias gramnegativas, por lo que es probable que uno haya evolucionado del otro. Debido a que el T3SS tiene una cantidad similar de componentes que un aparato flagelar (alrededor de 25 proteínas), es difícil determinar cuál evolucionó primero. Sin embargo, el sistema flagelar parece involucrar más proteínas en general, incluidos varios reguladores y chaperonas, por lo que se ha argumentado que los flagelos evolucionaron a partir de un T3SS. Sin embargo, también se ha sugerido que el flagelo pudo haber evolucionado primero o que las dos estructuras evolucionaron en paralelo. Los primeros organismos unicelulares' La necesidad de motilidad (movilidad) respalda que los flagelos más móviles serían seleccionados primero por la evolución, pero el T3SS que evoluciona del flagelo puede verse como una "evolución reductiva" y no recibe apoyo topológico de los árboles filogenéticos. La hipótesis de que las dos estructuras evolucionaron por separado a partir de un ancestro común explica las similitudes proteicas entre las dos estructuras, así como su diversidad funcional.
Flagelos y el debate del diseño inteligente
Algunos autores han argumentado que los flagelos no pueden haber evolucionado, asumiendo que solo pueden funcionar correctamente cuando todas las proteínas están en su lugar. En otras palabras, el aparato flagelar es "irreductiblemente complejo". Sin embargo, muchas proteínas se pueden eliminar o mutar y el flagelo aún funciona, aunque a veces con una eficiencia reducida. Además, con muchas proteínas exclusivas de algunas especies, la diversidad de la composición de los flagelos bacterianos fue mayor de lo esperado. Por tanto, el aparato flagelar es claramente muy flexible en términos evolutivos y perfectamente capaz de perder o ganar componentes proteicos. Por ejemplo, se han encontrado varias mutaciones que aumentan la motilidad de E. coli. La evidencia adicional de la evolución de los flagelos bacterianos incluye la existencia de flagelos vestigiales, formas intermedias de flagelos y patrones de similitudes entre las secuencias de proteínas flagelares, incluida la observación de que casi todas las proteínas flagelares centrales tienen homologías conocidas con proteínas no flagelares. Además, se han identificado varios procesos que juegan un papel importante en la evolución flagelar, incluido el autoensamblaje de subunidades repetitivas simples, la duplicación de genes con divergencia posterior, el reclutamiento de elementos de otros sistemas ('bricolage molecular') y la recombinación.
Arreglos flagelares
Diferentes especies de bacterias tienen diferentes números y arreglos de flagelos, nombrados usando el término tricho, del griego trichos que significa pelo.
- Monotrichous bacterias tales como Vibrio cholerae tener un solo polar flagellum.
- Amphitrichous las bacterias tienen un único flagellum en cada uno de los dos extremos opuestos (por ejemplo, Alcaligenes faecalis), sólo un flagellum opera a la vez, permitiendo que la bacteria revierta rápidamente el curso cambiando qué flagellum es activo.
- Lophotrichous bacterias ()lopho Griego combinando término significando crest o tuft) tienen múltiples flagella ubicada en el mismo lugar en la superficie bacteriana como Helicobacter pylori, que actúan en concierto para conducir la bacteria en una sola dirección. En muchos casos, las bases de la bandera múltiple están rodeadas por una región especializada de la membrana celular, llamada organela polar.
- Peritrichous bacterias tienen flagella proyectando en todas direcciones (por ejemplo, E. coli).
La rotación en sentido contrario a las agujas del reloj de un flagelo polar monotrichous empuja la célula hacia adelante con el flagelo detrás, como un sacacorchos que se mueve dentro del corcho. El agua a escala microscópica es muy viscosa, a diferencia del agua habitual.
Las espiroquetas, por el contrario, tienen flagelos llamados endoflagelos que surgen de los polos opuestos de la célula y están ubicados dentro del espacio periplásmico como se muestra al romper la membrana externa y también por microscopía de criotomografía electrónica. La rotación de los filamentos en relación con el cuerpo celular hace que toda la bacteria avance con un movimiento similar al de un sacacorchos, incluso a través de material lo suficientemente viscoso como para impedir el paso de bacterias normalmente flageladas.
En ciertas formas grandes de Selenomonas, más de 30 flagelos individuales están organizados fuera del cuerpo celular, enroscándose helicoidalmente entre sí para formar una estructura gruesa (fácilmente visible con el microscopio óptico) llamada & #34;fascículo".
En algunas Vibrio spp. (particularmente Vibrio parahaemolyticus) y bacterias relacionadas como Aeromonas, dos sistemas flagelares coexisten, utilizando diferentes conjuntos de genes y diferentes gradientes iónicos para obtener energía. Los flagelos polares se expresan constitutivamente y proporcionan motilidad en el fluido a granel, mientras que los flagelos laterales se expresan cuando los flagelos polares encuentran demasiada resistencia para girar. Estos proporcionan motilidad de enjambre en superficies o en fluidos viscosos.
Agrupar
La agrupación es un evento que puede ocurrir en células multiflageladas, uniendo los flagelos y haciendo que giren de manera coordinada.
Los flagelos son hélices levógiras y, cuando sus rotores los giran en sentido contrario a las agujas del reloj, pueden agruparse y girar juntos. Cuando los rotores invierten la dirección, girando así en el sentido de las agujas del reloj, el flagelo se desenrolla del haz. Esto puede hacer que la célula detenga su movimiento hacia adelante y, en su lugar, comience a retorcerse en su lugar, lo que se conoce como "tumbling". La caída da como resultado una reorientación estocástica de la celda, lo que hace que cambie la dirección de su natación hacia adelante.
No se sabe qué estímulos impulsan el cambio entre agrupamiento y volteo, pero el motor se adapta muy bien a diferentes señales. En el modelo que describe la quimiotaxis ("movimiento a propósito"), la rotación en el sentido de las agujas del reloj de un flagelo es suprimida por compuestos químicos favorables para la célula (por ejemplo, alimentos). Cuando se mueve en una dirección favorable, la concentración de tales atrayentes químicos aumenta y, por lo tanto, las volteretas se suprimen continuamente, lo que permite el movimiento hacia adelante; Del mismo modo, cuando la dirección de movimiento de la célula es desfavorable (por ejemplo, lejos de un atrayente químico), las caídas ya no se suprimen y ocurren con mucha más frecuencia, con la posibilidad de que la célula se reoriente en la dirección correcta.
Incluso si todos los flagelos giraran en el sentido de las agujas del reloj, a menudo no pueden formar un paquete debido a razones geométricas e hidrodinámicas.
Eucariótico




Terminología
Con el objetivo de enfatizar la distinción entre los flagelos bacterianos y los cilios y flagelos eucariotas, algunos autores intentaron reemplazar el nombre de estas dos estructuras eucariotas con "undulipodia" (p. ej., todos los artículos de Margulis desde la década de 1970) o "cilia" para ambos (p. ej., Hülsmann, 1992; Adl et al., 2012; la mayoría de los artículos de Cavalier-Smith), conservando "flagelos" por la estructura bacteriana. Sin embargo, el uso discriminatorio de los términos "cilia" y "flagelos" para los eucariotas adoptado en este artículo (ver § Flagella versus cilia a continuación) sigue siendo común (p. ej., Andersen et al., 1991; Leadbeater et al., 2000).
Estructura interna
El núcleo de un flagelo eucariótico, conocido como axonema, es un haz de nueve pares de microtúbulos fusionados conocidos como dobles que rodean dos microtúbulos centrales únicos (singletes). Este axonema 9+2 es característico del flagelo eucariota. En la base de un flagelo eucariótico hay un cuerpo basal, "blefaroplasto" o cinetosoma, que es el centro organizador de microtúbulos para los microtúbulos flagelares y tiene una longitud de unos 500 nanómetros. Los cuerpos basales son estructuralmente idénticos a los centríolos. El flagelo está encerrado dentro de la membrana plasmática de la célula, de modo que el interior del flagelo es accesible al citoplasma de la célula.
Además del axonema y el cuerpo basal, de morfología relativamente constante, otras estructuras internas del aparato flagelar son la zona de transición (donde se unen el axonema y el cuerpo basal) y el sistema radicular (estructuras microtubulares o fibrilares que se extienden desde los cuerpos basales en el citoplasma), más variables y útiles como indicadores de las relaciones filogenéticas de los eucariotas. Otras estructuras, menos frecuentes, son el bastón paraflagelar (o paraxial, paraxonemal), la fibra R y la fibra S. Para estructuras superficiales, ver más abajo.
Mecanismo
Cada uno de los 9 microtúbulos dobles externos extiende un par de brazos de dineína (un brazo 'interior' y otro 'externo') al microtúbulo adyacente; estos producen fuerza a través de la hidrólisis de ATP. El axonema flagelar también contiene radios radiales, complejos polipeptídicos que se extienden desde cada uno de los nueve dobletes de microtúbulos externos hacia el par central, con la "cabeza" del radio hacia adentro. Se cree que el radio radial está involucrado en la regulación del movimiento flagelar, aunque aún no se conocen su función exacta y su método de acción.
Flagelos versus cilios

Los patrones de latidos regulares de los cilios y flagelos eucariotas generan movimiento a nivel celular. Los ejemplos van desde la propulsión de células individuales, como la natación de los espermatozoides, hasta el transporte de fluidos a lo largo de una capa estacionaria de células, como en el tracto respiratorio.
Aunque los cilios y flagelos eucarióticos son en última instancia lo mismo, a veces se clasifican por su patrón de movimiento, una tradición anterior a que se conocieran sus estructuras. En el caso de los flagelos, el movimiento suele ser plano y ondulatorio, mientras que los cilios móviles suelen realizar un movimiento tridimensional más complicado con un golpe de potencia y recuperación. Otra forma tradicional de distinción es por el número de orgánulos 9+2 en la célula.
Transporte intraflagelar
El transporte intraflagelar, el proceso mediante el cual las subunidades axonémicas, los receptores transmembrana y otras proteínas se mueven hacia arriba y hacia abajo a lo largo del flagelo, es esencial para el correcto funcionamiento del flagelo, tanto en la motilidad como en la transducción de señales.
Evolución y ocurrencia
Los flagelos o cilios eucariotas, probablemente una característica ancestral, están muy extendidos en casi todos los grupos de eucariotas, como una condición relativamente perenne o como una etapa flagelada del ciclo de vida (por ejemplo, zoides, gametos, zoosporas, que pueden producirse de forma continua o continua). no).
La primera situación se encuentra en células especializadas de organismos multicelulares (p. ej., los coanocitos de las esponjas o el epitelio ciliado de los metazoos), como en los ciliados y muchos eucariotas con una "condición flagelada" (o "nivel de organización monadoide", véase Flagellata, un grupo artificial).
Las etapas flageladas del ciclo de vida se encuentran en muchos grupos, por ejemplo, muchas algas verdes (zoosporas y gametos masculinos), briófitas (gametos masculinos), pteridófitas (gametos masculinos), algunas gimnospermas (cícadas y Ginkgo, como gametos masculinos), diatomeas céntricas (gametos masculinos), algas pardas (zoosporas y gametos), oomicetos (zoosporas y gametos asexuales), hifoquítridos (zoosporas), laberíntulomicetos (zoosporas), algunos apicomplexanos (gametos), algunos radiolarios (probablemente gametos), foraminíferos (gametos), plasmodioforomicetos (zoosporas y gametos), mixogástridos (zoosporas), metazoos (gametos masculinos) y hongos quítridos (zoosporas y gametos).
Los flagelos o los cilios están completamente ausentes en algunos grupos, probablemente debido a una pérdida más que a una condición primitiva. La pérdida de cilios se produjo en las algas rojas, algunas algas verdes (Zygnematophyceae), las gimnospermas excepto las cícadas y Ginkgo, las angiospermas, las diatomeas pennadas, algunos apicomplejos, algunos amebozoos, en el esperma de algunos metazoos y en hongos (excepto quitridios).
Tipología
Se utilizan varios términos relacionados con flagelos o cilios para caracterizar a los eucariotas. Según las estructuras superficiales presentes, los flagelos pueden ser:
- whiplash flagella (= suave, acronematic flagella): sin pelos, por ejemplo, en Opisthokonta
- flagella peluda (= tinsel, flimmer, pleuronematic flagella): con pelos (= mastigonemes sensu lato), dividido en:
- con cabellos finos (= no-tubulares, o pelos simples): ocurre en Euglenophyceae, Dinoflagellata, algunos Haptophyceae (Pavlovales)
- con cabellos rígidos (= pelos tubulares, retronemes, mastigonemes sensu stricto), dividido en:
- pelos bipartitos: con dos regiones. Occurs in Cryptophyceae, Prasinophyceae, and some Heterokonta
- pelos tripartitos (= estraminipilosos): con tres regiones (una base, un eje tubular y uno o más pelos terminales). Occurs in most Heterokonta
- stichonematic flagella: con una sola fila de cabellos
- pantonematic flagella: con dos filas de cabellos
- acronematic: flagella con un solo, terminal mastigoneme o flagellar hair (por ejemplo, bodonids); algunos autores utilizan el término como sinónimo de whiplash
- con escalas: por ejemplo, Prasinophyceae
- con espinas: por ejemplo, algunas algas marrones
- con membrana ondulante: por ejemplo, algunos kinetoplastoides, algunos parabasálidos
- con proboscis (protrusión parecida a la de la célula): por ejemplo, apusomonads, algunos bodonoides
Según el número de flagelos, las células pueden ser: (recordando que algunos autores usan "ciliadas" en lugar de "flageladas")
- uniflagellated: por ejemplo, la mayoría de Opisthokonta
- biflagellated: por ejemplo, todos Dinoflagellata, los gametos de Charophyceae, de la mayoría de los biófitos y de algunos metazoanos
- triflagellated: por ejemplo, los gametos de algunos Foraminifera
- quadriflagellated: por ejemplo, algunos Prasinophyceae, Collodictyonidae
- octoflagellated: e.g., some Diplomonada, some Prasinophyceae
- multiflagellated: por ejemplo, Opalinata, Ciliophora, Stephanopogon, Parabasalida, Hemimastigophora, Caryoblastea, Multicilia, los gametos (o zoides) de Oedogoniales (Chlorophyta), algunos pteridophytes y algunos gimnasiosperms
Según el lugar de inserción de los flagelos:
- opisthokont: células con flagella insertadas posteriormente, por ejemplo, en Opisthokonta (Vischer, 1945). En Haptophyceae, la flagella se inserta lateralmente a terminal, pero se dirigen posteriormente durante la natación rápida.
- akrokont: células con flagella insertadas apically
- subakrokont: células con flagella insertadas subapicamente
- pleurokont: células con flagella insertada lateralmente
Según el patrón de golpes:
- gliding: un flagellum que recorre el sustrato
- heterodinámica: flagelo con diferentes patrones de golpiza (generalmente con un flagelo que funciona en la captura de alimentos y el otro funcionamiento en el gliding, el anclaje, la propulsión o el "steering")
- isodinámica: flagella golpeando con los mismos patrones
Otros términos relacionados con el tipo flagelar:
- isokont: celdas con flagella de igual longitud. También se utilizó anteriormente para referirse a la clorophyta
- anisokont: células con flagelo de longitud desigual, por ejemplo, algunas Euglenophyceae y Prasinophyceae
- heterokont: término introducido por Lutero (1899) para referirse a la Xanthophyceae, debido al par de flagella de longitud desigual. Ha tomado un significado específico al referirse a las células con un flagelo estraminoso anterior (con mastigomas tripartitos, en una o dos filas) y un flagellón generalmente liso posterior. También se utiliza para referirse al taxón Heterokonta
- stephanokont: células con una corona de flagella cerca de su extremo anterior, por ejemplo, los gametos y esporas de Oedogoniales, las esporas de algunos Bryopsidales. Término introducido por Blackman " Tansley (1902) para referirse a los Oedogoniales
- akont: células sin flagella. También se utilizó para referirse a grupos taxonómicos, como Aconta o Akonta: el Zygnematophyceae y Bacillariophyceae (Oltmanns, 1904), o el Rhodophyceae (Christensen, 1962)
Arqueal
El archaellum que poseen algunas especies de Archaea es superficialmente similar al flagelo bacteriano; en la década de 1980, se pensó que eran homólogos sobre la base de la morfología y el comportamiento generales. Tanto los flagelos como las archaellas consisten en filamentos que se extienden fuera de la célula y giran para impulsar la célula. Archaeal flagelos tienen una estructura única que carece de un canal central. Al igual que las pilinas bacterianas de tipo IV, las proteínas arqueales (arqueelinas) se elaboran con péptidos señal de clase 3 y son procesadas por una enzima similar a la prepilina peptidasa de tipo IV. Las arquelinas se modifican normalmente mediante la adición de glucanos unidos a N que son necesarios para un ensamblaje o funcionamiento adecuados.
Los descubrimientos en la década de 1990 revelaron numerosas diferencias detalladas entre los flagelos arqueales y bacterianos. Éstos incluyen:
- La rotación bacteriana de la bandera es alimentada por la fuerza motriz protona – un flujo de iones H+ o ocasionalmente por la fuerza sodio-motiva – un flujo de iones Na+; la rotación de la bandera arqueal es alimentada por ATP.
- Mientras que las células bacterianas suelen tener muchos filamentos flagelador, cada uno de los cuales gira independientemente, el flagelo arqueal se compone de un paquete de muchos filamentos que giran como una sola asamblea.
- La flagelación bacteriana crece por la adición de subunidades de bandera en la punta; la flagella arqueal crece por la adición de subunidades a la base.
- La flagelo bacteriano es más gruesa que la arquela, y el filamento bacteriano tiene un "tubo" hueco lo suficientemente grande dentro que las subunidades banderales pueden fluir por el interior del filamento y se añaden en la punta; el arquelio es demasiado delgado (12-15 nm) para permitir esto.
- Muchos componentes de flagelo bacteriano comparten la semejanza de secuencia a componentes de los sistemas de secreción tipo III, pero los componentes de flagelo bacteriano y arquela no comparten la semejanza de secuencia. En cambio, algunos componentes de la secuencia de acciones de la arquela y la similitud morfológica con componentes de pili tipo IV, que se montan a través de la acción de los sistemas de secreción tipo II (la nomenclatura de los sistemas de pili y proteína no es consistente).
Estas diferencias podrían significar que los flagelos y archaellas bacterianos podrían ser un caso clásico de analogía biológica, o evolución convergente, en lugar de homología. Sin embargo, en comparación con las décadas de estudio bien publicitado de los flagelos bacterianos (por ejemplo, por Howard Berg), las archaella han comenzado a atraer la atención científica recientemente.
Imágenes adicionales
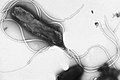
Múltiple flagella en el arranjo de lophotrichous en la superficie de Helicobacter pylori

Modelo físico de un flagelo bacteriano


Contenido relacionado
Buitre del Nuevo Mundo
Pterosaurio
Trogón