
Enlace peptídico
En química orgánica, un enlace peptídico es un tipo de enlace químico covalente de amida que une dos alfa-aminoácidos consecutivos de C1 (carbono número uno) de un alfa-aminoácido y N2 (nitrógeno número dos) de otro, a lo largo de un péptido o proteína. cadena.
También puede llamarse enlace eupéptido para distinguirlo de un enlace isopéptido, que es otro tipo de enlace amida entre dos aminoácidos.
Síntesis
Cuando dos aminoácidos forman un dipéptido a través de un enlace peptídico, es un tipo de reacción de condensación. En este tipo de condensación, dos aminoácidos se acercan entre sí, con el resto de ácido carboxílico de cadena no lateral (C1) de uno acercándose al resto amino de cadena no lateral (N2) del otro. Uno pierde hidrógeno y oxígeno de su grupo carboxilo (COOH) y el otro pierde hidrógeno de su grupo amino (NH 2). Esta reacción produce una molécula de agua (H 2 O) y dos aminoácidos unidos por un enlace peptídico (−CO−NH−). Los dos aminoácidos unidos se denominan dipéptido.
El enlace amida se sintetiza cuando el grupo carboxilo de una molécula de aminoácido reacciona con el grupo amino de la otra molécula de aminoácido, provocando la liberación de una molécula de agua (H 2 O), por lo que el proceso es una reacción de síntesis por deshidratación.
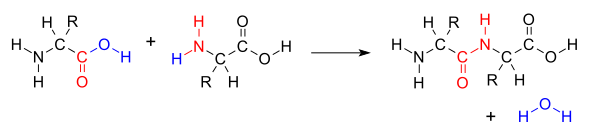 La condensación por deshidratación de dos aminoácidos para formar un enlace peptídico (rojo) con expulsión de agua (azul)
La condensación por deshidratación de dos aminoácidos para formar un enlace peptídico (rojo) con expulsión de agua (azul)
La formación del enlace peptídico consume energía que, en los organismos, se deriva del ATP. Los péptidos y las proteínas son cadenas de aminoácidos que se mantienen unidas por enlaces peptídicos (ya veces por unos pocos enlaces isopeptídicos). Los organismos utilizan enzimas para producir péptidos no ribosómicos y ribosomas para producir proteínas a través de reacciones que difieren en detalles de la síntesis por deshidratación.
Algunos péptidos, como la alfa-amanitina, se denominan péptidos ribosómicos porque los fabrican los ribosomas, pero muchos son péptidos no ribosómicos porque los sintetizan enzimas especializadas en lugar de ribosomas. Por ejemplo, el tripéptido glutatión se sintetiza en dos pasos a partir de aminoácidos libres, mediante dos enzimas: glutamato-cisteína ligasa (forma un enlace isopeptídico, que no es un enlace peptídico) y glutatión sintetasa (forma un enlace peptídico).
Degradación
Un enlace peptídico se puede romper por hidrólisis (la adición de agua). La hidrólisis de los enlaces peptídicos en el agua libera 8 a 16 kJ/mol (2 a 4 kcal/mol) de energía de Gibbs. Este proceso es extremadamente lento, con una vida media a 25 °C de entre 350 y 600 años por bono.
En los organismos vivos, el proceso normalmente es catalizado por enzimas conocidas como peptidasas o proteasas, aunque hay informes de hidrólisis de enlaces peptídicos causada por tensión conformacional cuando el péptido/proteína se pliega en la estructura nativa. Este proceso no enzimático no es acelerado por la estabilización del estado de transición, sino más bien por la desestabilización del estado fundamental.
Espectros
La longitud de onda de absorción de un enlace peptídico es de 190 a 230 nm, lo que lo hace particularmente susceptible a la radiación ultravioleta.
Isómeros cis/trans del grupo peptídico
La deslocalización significativa del par solitario de electrones en el átomo de nitrógeno le da al grupo un carácter de doble enlace parcial. El doble enlace parcial hace que el grupo amida sea plano, ya sea en los isómeros cis o trans. En el estado desplegado de las proteínas, los grupos peptídicos están libres para isomerizarse y adoptar ambos isómeros; sin embargo, en el estado plegado, solo se adopta un solo isómero en cada posición (con raras excepciones). La forma trans se prefiere abrumadoramente en la mayoría de los enlaces peptídicos (proporción de aproximadamente 1000:1 en poblaciones trans:cis). Sin embargo, los grupos peptídicos X-Pro tienden a tener una proporción aproximada de 30:1, presumiblemente porque la simetría entre los átomos C y C de la prolina hace que los isómeros cis y trans sean casi iguales en energía, véase la figura.
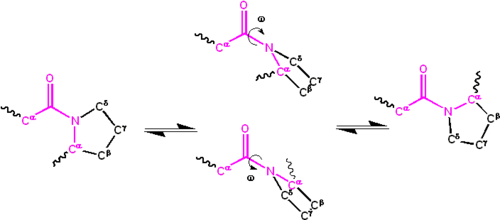 Isomerización de un enlace peptídico X-Pro. Los isómeros cis y trans están en el extremo izquierdo y derecho, respectivamente, separados por los estados de transición.
Isomerización de un enlace peptídico X-Pro. Los isómeros cis y trans están en el extremo izquierdo y derecho, respectivamente, separados por los estados de transición.
Se denota el ángulo diedro asociado con el grupo peptídico (definido por los cuatro átomos C –C'–N–C) 




El plegamiento de proteínas conformacionales suele ser mucho más rápido (típicamente 10 a 100 ms) que la isomerización cis-trans (10 a 100 s). Un isómero no nativo de algunos grupos peptídicos puede alterar significativamente el plegamiento conformacional, ya sea ralentizándolo o impidiendo que ocurra hasta que se alcance el isómero nativo. Sin embargo, no todos los grupos peptídicos tienen el mismo efecto sobre el plegamiento; los isómeros no nativos de otros grupos peptídicos pueden no afectar en absoluto al plegamiento.
Reacciones químicas
Debido a su estabilización por resonancia, el enlace peptídico es relativamente poco reactivo en condiciones fisiológicas, incluso menos que compuestos similares como los ésteres. Sin embargo, los enlaces peptídicos pueden sufrir reacciones químicas, generalmente a través de un ataque de un átomo electronegativo en el carbono del carbonilo, rompiendo el doble enlace del carbonilo y formando un intermedio tetraédrico. Esta es la vía seguida en la proteólisis y, más generalmente, en las reacciones de intercambio de N-O acilo como las de las inteínas. Cuando el grupo funcional que ataca el enlace peptídico es un tiol, un hidroxilo o una amina, la molécula resultante puede denominarse ciclol o, más específicamente, tiaciclol, oxaciclol o azaciclol, respectivamente.
Contenido relacionado
Higroscopia
Radio atómico
Hidrogenación