
Dominancia (genética)
En genética, la dominancia es el fenómeno de una variante (alelo) de un gen en un cromosoma que enmascara o anula el efecto de una variante diferente del mismo gen en la otra copia del cromosoma. La primera variante se denomina dominante y la segunda recesiva. Este estado de tener dos variantes diferentes del mismo gen en cada cromosoma es causado originalmente por una mutación en uno de los genes, ya sea nuevo ( de novo ) o heredado. Los términos autosómico dominante o autosómico recesivose utilizan para describir variantes genéticas en cromosomas no sexuales (autosomas) y sus características asociadas, mientras que las de los cromosomas sexuales (alosomas) se denominan dominantes ligadas al X, recesivas ligadas al X o ligadas al Y; estos tienen un patrón de herencia y presentación que depende del sexo tanto del padre como del hijo (ver Vinculación sexual). Dado que solo hay una copia del cromosoma Y, los rasgos ligados a Y no pueden ser dominantes ni recesivos. Además, existen otras formas de dominancia, como la dominancia incompleta, en la que una variante genética tiene un efecto parcial en comparación con cuando está presente en ambos cromosomas, y la codominancia, en la que diferentes variantes en cada cromosoma muestran sus rasgos asociados.
La dominancia es un concepto clave en la herencia mendeliana y la genética clásica. Se utilizan letras y cuadros de Punnett para demostrar los principios de dominancia en la enseñanza, y el uso de letras mayúsculas para los alelos dominantes y minúsculas para los alelos recesivos es una convención muy seguida. Un ejemplo clásico de dominancia es la herencia de la forma de la semilla en los guisantes. Los guisantes pueden ser redondos, asociados con el alelo R, o arrugados, asociados con el alelo r. En este caso, son posibles tres combinaciones de alelos (genotipos): RR, Rr y rr. Los individuos RR (homocigotos) tienen guisantes redondos y los individuos rr (homocigotos) tienen guisantes rugosos. EnRr (heterocigotos), el alelo R enmascara la presencia del alelo r, por lo que estos individuos también tienen guisantes redondos. Por lo tanto, el alelo R es dominante sobre el alelo r, y el alelo r es recesivo con respecto al alelo R.
La dominancia no es inherente a un alelo o sus rasgos (fenotipo). Es un efecto estrictamente relativo entre dos alelos de un gen dado de cualquier función; un alelo puede ser dominante sobre un segundo alelo del mismo gen, recesivo para un tercero y codominante con un cuarto. Además, un alelo puede ser dominante para un rasgo pero no para otros.
La dominancia difiere de la epistasis, el fenómeno de un alelo de un gen que enmascara el efecto de los alelos de un gen diferente.
Fondo
El concepto de dominación fue introducido por Gregor Johann Mendel. Aunque Mendel, "El padre de la genética", utilizó el término por primera vez en la década de 1860, no fue muy conocido hasta principios del siglo XX. Mendel observó que, para una variedad de características de los guisantes de jardín que tienen que ver con la apariencia de las semillas, las vainas de las semillas y las plantas, había dos fenotipos discretos, como semillas redondas frente a rugosas, semillas amarillas frente a verdes, flores rojas frente a blancas o plantas altas versus bajas. Cuando se criaron por separado, las plantas siempre produjeron los mismos fenotipos, generación tras generación. Sin embargo, cuando se cruzaron (cruzaron) líneas con diferentes fenotipos, uno y solo uno de los fenotipos parentales apareció en la descendencia (verde, redondo, rojo o alto). Sin embargo, cuando se cruzaron estas plantas híbridas, las plantas descendientes mostraron los dos fenotipos originales, en una proporción característica de 3:1, siendo el fenotipo más común el de las plantas híbridas progenitoras. Mendel razonó que cada padre en el primer cruce era un homocigoto para diferentes alelos (un padre AA y el otro padre aa), que cada uno aportaba un alelo a la descendencia, con el resultado de que todos estos híbridos eran heterocigotos (Aa), y que uno de los dos alelos en el cruce híbrido dominaba la expresión del otro: A enmascarado a. El cruce final entre dos heterocigotos (Aa X Aa) produciría descendencia AA, Aa y aa en una proporción de genotipo 1:2:1 con las dos primeras clases mostrando el fenotipo (A) y la última mostrando el fenotipo (a)., produciendo así la proporción de fenotipo 3:1. siendo el fenotipo más común el de las plantas híbridas parentales. Mendel razonó que cada padre en el primer cruce era un homocigoto para diferentes alelos (un padre AA y el otro padre aa), que cada uno aportaba un alelo a la descendencia, con el resultado de que todos estos híbridos eran heterocigotos (Aa), y que uno de los dos alelos en el cruce híbrido dominaba la expresión del otro: A enmascarado a. El cruce final entre dos heterocigotos (Aa X Aa) produciría descendencia AA, Aa y aa en una proporción de genotipo 1:2:1 con las dos primeras clases mostrando el fenotipo (A) y la última mostrando el fenotipo (a)., produciendo así la proporción de fenotipo 3:1. siendo el fenotipo más común el de las plantas híbridas parentales. Mendel razonó que cada padre en el primer cruce era un homocigoto para diferentes alelos (un padre AA y el otro padre aa), que cada uno aportaba un alelo a la descendencia, con el resultado de que todos estos híbridos eran heterocigotos (Aa), y que uno de los dos alelos en el cruce híbrido dominaba la expresión del otro: A enmascarado a. El cruce final entre dos heterocigotos (Aa X Aa) produciría descendencia AA, Aa y aa en una proporción de genotipo 1:2:1 con las dos primeras clases mostrando el fenotipo (A) y la última mostrando el fenotipo (a)., produciendo así la proporción de fenotipo 3:1. que cada uno aportó un alelo a la descendencia, con el resultado de que todos estos híbridos eran heterocigotos (Aa), y que uno de los dos alelos en el cruce híbrido dominaba la expresión del otro: A enmascarado a. El cruce final entre dos heterocigotos (Aa X Aa) produciría descendencia AA, Aa y aa en una proporción de genotipo 1:2:1 con las dos primeras clases mostrando el fenotipo (A) y la última mostrando el fenotipo (a)., produciendo así la proporción de fenotipo 3:1. que cada uno aportó un alelo a la descendencia, con el resultado de que todos estos híbridos eran heterocigotos (Aa), y que uno de los dos alelos en el cruce híbrido dominaba la expresión del otro: A enmascarado a. El cruce final entre dos heterocigotos (Aa X Aa) produciría descendencia AA, Aa y aa en una proporción de genotipo 1:2:1 con las dos primeras clases mostrando el fenotipo (A) y la última mostrando el fenotipo (a)., produciendo así la proporción de fenotipo 3:1.
Mendel no usó los términos gen, alelo, fenotipo, genotipo, homocigoto y heterocigoto, todos los cuales se introdujeron más tarde. Introdujo la notación de letras mayúsculas y minúsculas para alelos dominantes y recesivos, respectivamente, todavía en uso en la actualidad.
En 1928, el genetista de poblaciones británico Ronald Fisher propuso que la dominancia actuaba en base a la selección natural mediante la contribución de genes modificadores. En 1929, el genetista estadounidense Sewall Wright respondió afirmando que la dominancia es simplemente una consecuencia fisiológica de las vías metabólicas y la necesidad relativa del gen involucrado. La explicación de Wright se convirtió en un hecho establecido en genética y el debate terminó en gran medida. Sin embargo, algunos rasgos pueden tener su dominancia influenciada por mecanismos evolutivos.
Cromosomas, genes y alelos
La mayoría de los animales y algunas plantas tienen cromosomas emparejados y se describen como diploides. Tienen dos versiones de cada cromosoma, una aportada por el óvulo de la madre y la otra por el espermatozoide del padre, conocidos como gametos, descritos como haploides y creados a través de la meiosis. Estos gametos luego se fusionan durante la fertilización durante la reproducción sexual, en un nuevo cigoto de una sola célula, que se divide varias veces, dando como resultado un nuevo organismo con la misma cantidad de pares de cromosomas en cada célula (no gameto) que sus padres.
Cada cromosoma de un par coincidente (homólogo) es estructuralmente similar al otro y tiene una secuencia de ADN muy similar (loci, locus singular). El ADN en cada cromosoma funciona como una serie de genes discretos que influyen en varios rasgos. Así, cada gen también tiene un homólogo correspondiente, que puede existir en diferentes versiones llamadas alelos. Los alelos en el mismo locus en los dos cromosomas homólogos pueden ser idénticos o diferentes.
El tipo de sangre de un humano está determinado por un gen que crea un tipo de sangre A, B, AB u O y está ubicado en el brazo largo del cromosoma nueve. Hay tres alelos diferentes que podrían estar presentes en este locus, pero solo dos pueden estar presentes en cualquier individuo, uno heredado de su madre y otro de su padre.
Si dos alelos de un gen dado son idénticos, el organismo se denomina homocigoto y se dice que es homocigoto con respecto a ese gen; si en cambio los dos alelos son diferentes, el organismo es heterocigoto y es heterocigoto. La composición genética de un organismo, ya sea en un solo locus o en todos sus genes colectivamente, se denomina genotipo. El genotipo de un organismo, directa e indirectamente, afecta sus rasgos moleculares, físicos y otros, que individual o colectivamente se denominan fenotipo. En loci de genes heterocigotos, los dos alelos interactúan para producir el fenotipo.
Tipos de Dominancia
Dominio completo
En dominancia completa, el efecto de un alelo en un genotipo heterocigoto enmascara por completo el efecto del otro. El alelo que enmascara se considera dominante al otro alelo, y el alelo enmascarado se considera recesivo. La dominancia completa en el fenotipo de un heterocigoto es indistinguible del fenotipo de un homocigoto dominante.
Un ejemplo clásico de dominancia completa es la herencia de la forma de la semilla (forma de guisante) en los guisantes. Los guisantes pueden ser redondos (asociados con el alelo R ) o arrugados (asociados con el alelo r ). En este caso, son posibles tres combinaciones de alelos (genotipos): RR, rr, Rr. RR y rr son homocigotos y Rr es heterocigoto. Los individuos RR tienen guisantes redondos y los individuos rr tienen guisantes rugosos. En los individuos Rr, el alelo R enmascara la presencia del alelo r, por lo que estos individuos también tienen guisantes redondos. Por lo tanto, el alelo R es completamente dominante al alelor, y el alelo r es recesivo al alelo R.
Dominancia incompleta
Dominancia incompleta (también llamada dominancia parcial, semidominancia o herencia intermedia )) ocurre cuando el fenotipo del genotipo heterocigoto es distinto y a menudo intermedio a los fenotipos de los genotipos homocigotos. El resultado fenotípico a menudo aparece como una forma combinada de características en estado heterocigoto. Por ejemplo, el color de la flor boca de dragón es homocigoto para el rojo o el blanco. Cuando la flor homocigota roja se combina con la flor homocigota blanca, el resultado produce una flor de boca de dragón rosa. La boca de dragón rosa es el resultado de una dominancia incompleta. Un tipo similar de dominancia incompleta se encuentra en la planta de las cuatro en punto, en la que se produce un color rosa cuando se cruzan padres de raza pura de flores blancas y rojas. En genética cuantitativa, donde los fenotipos se miden y tratan numéricamente, si el fenotipo de un heterocigoto está exactamente entre (numéricamente) el de los dos homocigotos,no hay dominio en absoluto, es decir, el dominio existe sólo cuando la medida del fenotipo del heterocigoto se encuentra más cerca de un homocigoto que del otro.
Cuando las plantas de la generación F 1 se autopolinizan, la relación fenotípica y genotípica de la generación F 2 será de 1:2:1 (Rojo:Rosa:Blanco).
Ver hipótesis de dominancia parcial.
Co-dominancia
La codominancia ocurre cuando las contribuciones de ambos alelos son visibles en el fenotipo y ningún alelo enmascara al otro.
Por ejemplo, en el sistema de grupos sanguíneos ABO, las modificaciones químicas de una glicoproteína (el antígeno H) en la superficie de las células sanguíneas están controladas por tres alelos, dos de los cuales son co-dominantes entre sí ( I, I ) y dominantes sobre la i recesiva en el lugar geométrico ABO. Los alelos I e I producen diferentes modificaciones. La enzima codificada por I agrega una N-acetilgalactosamina a un antígeno H unido a la membrana. La enzima I añade una galactosa. El alelo i no produce modificación. Por lo tanto, los alelos I e I son dominantes para i ( I Iy los individuos I i ambos tienen sangre tipo A, y los individuos I I y I i ambos tienen sangre tipo B), pero los individuos I I tienen ambas modificaciones en sus células sanguíneas y, por lo tanto, tienen sangre tipo AB, por lo que los alelos I e I se denominan ser co-dominante.
Otro ejemplo ocurre en el locus del componente beta-globina de la hemoglobina, donde los tres fenotipos moleculares de Hb /Hb, Hb /Hb y Hb /Hb son todos distinguibles por electroforesis de proteínas. (La condición médica producida por el genotipo heterocigoto se denomina rasgo de células falciformes y es una condición más leve que se distingue de la anemia de células falciformes, por lo que los alelos muestran una dominancia incompleta con respecto a la anemia, véase más arriba). Para la mayoría de los loci de genes a nivel molecular, ambos alelos se expresan de forma codominante, porque ambos se transcriben en ARN.
La codominancia, donde los productos alélicos coexisten en el fenotipo, es diferente de la dominancia incompleta, donde la interacción cuantitativa de los productos alelos produce un fenotipo intermedio. Por ejemplo, en codominancia, una flor homocigota roja y una flor homocigota blanca producirán descendencia con manchas rojas y blancas. Cuando las plantas de la generación F1 se autopolinizan, la relación fenotípica y genotípica de la generación F2 será de 1:2:1 (Rojo:Manchado:Blanco). Estas proporciones son las mismas que las de la dominancia incompleta. Nuevamente, esta terminología clásica es inapropiada; en realidad, no debería decirse que tales casos muestren dominancia en absoluto.
Abordar conceptos erróneos comunes
La dominancia se relaciona con la relación entre dos versiones de un gen. Un rasgo dominante generalmente se corresponde con los patrones de herencia que se pueden ver en los cuadros de Punnett. Si un individuo tiene dos versiones de un gen, el gen que se observa con frecuencia en generaciones posteriores se considera "dominante".
En genética, hay algunos conceptos erróneos que son bastante comunes. Se piensa que un rasgo dominante es "más fuerte" y "supera" a un rasgo recesivo. También se supone que los rasgos dominantes tienen más probabilidades de ser heredados y más frecuentes en una población. La idea de que los rasgos dominantes sean masculinos o masculinos también es una suposición común en el mundo de la genética. Cualquier biólogo experimentado sabe que estas ideas no son fácticas, pero para un principiante estas ideas se destacan. Pero, si estas ideas no son fácticas, ¿por qué surgen? Las diferentes ideas de dominancia cuando se trata de genética se derivan de definiciones comúnmente conocidas de la palabra dominancia en un entorno no genético. Se piensa que la dominancia es controladora, fuerte y poderosa, lo que le da a la dominancia genética este estigma defectuoso.
La dominancia no determina si un alelo es perjudicial, neutral o ventajoso. Sin embargo, la selección debe operar sobre los genes indirectamente a través de los fenotipos y la dominancia afecta la exposición de los alelos en los fenotipos, por lo tanto, la tasa de cambio en las frecuencias alélicas bajo selección. Los alelos recesivos deletéreos pueden persistir en una población a bajas frecuencias, con la mayoría de las copias en heterocigotos, sin costo alguno para esos individuos. Estos raros recesivos son la base de muchos trastornos genéticos hereditarios.
Nomenclatura
En genética, los símbolos comenzaron como marcadores de posición algebraicos. Cuando un alelo es dominante sobre otro, la convención más antigua es simbolizar el alelo dominante con una letra mayúscula. Al alelo recesivo se le asigna la misma letra en minúsculas. En el ejemplo del guisante, una vez que se conoce la relación de dominancia entre los dos alelos, es posible designar el alelo dominante que produce una forma redonda con un símbolo de letra mayúscula R, y el alelo recesivo que produce una forma arrugada con una letra minúscula. caso símbolo r. Los genotipos homocigoto dominante, heterocigoto y homocigoto recesivo se escriben RR, Rr y rr, respectivamente. También sería posible designar los dos alelos como Wy w, y los tres genotipos WW, Ww y ww, los dos primeros de los cuales produjeron guisantes redondos y el tercero guisantes rugosos. La elección de " R " o " W " como símbolo para el alelo dominante no prejuzga si el alelo que causa el fenotipo "redondo" o "arrugado" cuando el homocigoto es el dominante.
Un gen puede tener varios alelos. Cada alelo está simbolizado por el símbolo del locus seguido de un superíndice único. En muchas especies, el alelo más común en la población salvaje se denomina alelo de tipo salvaje. Se simboliza con un carácter + como superíndice. Otros alelos son dominantes o recesivos al alelo de tipo salvaje. Para los alelos recesivos, el símbolo del locus está en minúsculas. Para los alelos con cualquier grado de dominancia sobre el alelo de tipo salvaje, la primera letra del símbolo del locus está en mayúsculas. Por ejemplo, estos son algunos de los alelos en el locus a del ratón de laboratorio, Mus musculus : A, amarillo dominante; a, tipo salvaje; y un, negro y fuego. el unel alelo es recesivo con respecto al alelo de tipo salvaje, y el alelo A es codominante con el alelo de tipo salvaje. El alelo A también es codominante con el alelo a, pero muestra que la relación está más allá de los límites de las reglas para la nomenclatura genética del ratón.
Las reglas de la nomenclatura genética han evolucionado a medida que la genética se ha vuelto más compleja. Los comités han estandarizado las reglas para algunas especies, pero no para todas. Las reglas para una especie pueden diferir un poco de las reglas para una especie diferente.
Relación con otros conceptos genéticos
Múltiples alelos
Aunque cualquier individuo de un organismo diploide tiene como máximo dos alelos diferentes en cualquier locus (salvo aneuploidías), la mayoría de los genes existen en un gran número de versiones alélicas en la población como un todo. Si los alelos tienen diferentes efectos sobre el fenotipo, a veces sus relaciones de dominancia pueden describirse como una serie.
Por ejemplo, el color del pelaje de los gatos domésticos se ve afectado por una serie de alelos del gen TYR (que codifica la enzima tirosinasa). Los alelos C, c, c y c (a todo color, birmano, siamés y albino, respectivamente) producen diferentes niveles de pigmento y, por lo tanto, diferentes niveles de dilución del color. El alelo C (a todo color) es completamente dominante sobre los tres últimos y el alelo c (albino) es completamente recesivo con respecto a los tres primeros.
Dominancia autosómica versus dominancia ligada al sexo
En los humanos y otras especies de mamíferos, el sexo está determinado por dos cromosomas sexuales llamados cromosoma X y cromosoma Y. Las hembras humanas son típicamente XX ; los machos son típicamente XY. Los pares restantes de cromosomas se encuentran en ambos sexos y se denominan autosomas; los rasgos genéticos debidos a los loci en estos cromosomas se describen como autosómicos y pueden ser dominantes o recesivos. Los rasgos genéticos en los cromosomas X e Y se denominan ligados al sexo porque están ligados a los cromosomas sexuales, no porque sean característicos de un sexo o del otro. En la práctica, el término casi siempre se refiere a XLos rasgos vinculados y muchos de esos rasgos (como la deficiencia de la visión del color rojo-verde) no se ven afectados por el sexo. Las hembras tienen dos copias de cada locus genético que se encuentra en el cromosoma X, al igual que los autosomas, y se aplican las mismas relaciones de dominancia. Los machos, sin embargo, tienen solo una copia de cada locus del gen del cromosoma X y se describen como hemicigotos para estos genes. El cromosoma Y es mucho más pequeño que el X., y contiene un conjunto mucho más pequeño de genes, incluidos, entre otros, aquellos que influyen en la "masculinidad", como el gen SRY para el factor determinante de los testículos. Las reglas de dominancia para los loci de genes ligados al sexo están determinadas por su comportamiento en la mujer: debido a que el hombre tiene solo un alelo (excepto en el caso de ciertos tipos de aneuploidía del cromosoma Y), ese alelo siempre se expresa independientemente de si es dominante o no. recesivo. Las aves tienen cromosomas sexuales opuestos: los machos tienen cromosomas ZZ y las hembras ZW. Sin embargo, la herencia de rasgos recuerda lo contrario al sistema XY; Los pinzones cebra machos pueden portar el gen de coloración blanca en uno de los dos cromosomas Z, pero las hembras siempre desarrollan coloración blanca. Los saltamontes tienen el sistema XO. Las mujeres tienen XX, pero los hombres solo X. No hay cromosoma Y en absoluto.
Epistasis
La epistasis [" epi + stasis = sentarse en la parte superior"] es una interacción entre alelos en dos loci de genes diferentes que afectan un solo rasgo, que a veces puede parecerse a una interacción de dominancia entre dos alelos diferentes en el mismo locus. La epistasis modifica la relación característica 9:3:3:1 esperada para dos genes no epistáticos. Para dos loci, se reconocen 14 clases de interacciones epistáticas. Como ejemplo de epistasis recesiva, un locus genético puede determinar si el pigmento de una flor es amarillo ( AA o Aa ) o verde ( aa ), mientras que otro locus determina si se produce el pigmento ( BBo Bb ) o no ( bb ). En una planta bb, las flores serán blancas, independientemente del genotipo del otro locus como AA, Aa o aa. La combinación bb no es dominante para el alelo A : más bien, el gen B muestra epistasis recesiva al gen A, porque el locus B cuando es homocigoto para el alelo recesivo ( bb ) suprime la expresión fenotípica del locus A. En un cruce entre dos AaBbplantas, esto produce una proporción característica de 9:3:4, en este caso de flores amarillas: verdes: blancas.
En la epistasis dominante, un locus de un gen puede determinar un pigmento amarillo o verde como en el ejemplo anterior: AA y Aa son amarillos y aa son verdes. Un segundo locus determina si se produce un precursor de pigmento ( dd ) o no ( DD o Dd ). Aquí, en una planta DD o Dd, las flores serán incoloras independientemente del genotipo en el locus A, debido al efecto epistático del alelo D dominante. Así, en un cruce entre dos plantas AaDd, 3/4 de las plantas serán incoloras, y los fenotipos amarillo y verde se expresan solo endd plantas. Esto produce una proporción característica de 12:3:1 de plantas blancas: amarillas: verdes.
La epistasis suplementaria ocurre cuando dos loci afectan el mismo fenotipo. Por ejemplo, si el color del pigmento es producido por CC o Cc pero no por cc, y por DD o Dd pero no por dd, entonces el pigmento no se produce en ninguna combinación genotípica con cc o dd. Es decir, ambos loci deben tener al menos un alelo dominante para producir el fenotipo. Esto produce una proporción característica de 9:7 de plantas pigmentadas a no pigmentadas. La epistasis complementaria en cambio produce una planta no pigmentada si y solo si el genotipo es cc y dd, y la relación característica es de 15:1 entre plantas pigmentadas y no pigmentadas.
La genética clásica consideraba interacciones epistáticas entre dos genes a la vez. Ahora es evidente a partir de la genética molecular que todos los loci de genes están involucrados en interacciones complejas con muchos otros genes (por ejemplo, las vías metabólicas pueden involucrar decenas de genes), y que esto crea interacciones epistáticas que son mucho más complejas que los modelos clásicos de dos locus..
Principio de Hardy-Weinberg (estimación de la frecuencia de la portadora)
La frecuencia del estado heterocigoto (que es el estado portador de un rasgo recesivo) se puede estimar utilizando la fórmula de Hardy-Weinberg:{displaystyle p^{2}+2pq+q^{2}=1}

Esta fórmula se aplica a un gen con exactamente dos alelos y relaciona las frecuencias de esos alelos en una población grande con las frecuencias de sus tres genotipos en esa población.
Por ejemplo, si p es la frecuencia del alelo A y q es la frecuencia del alelo a, entonces los términos p, 2, pq y q son las frecuencias de los genotipos AA, Aa y aa respectivamente. Dado que el gen tiene solo dos alelos, todos los alelos deben ser A o a y p + q = 1. Ahora, si A es completamente dominante a entonces la frecuencia del genotipo portador Aano se puede observar directamente (ya que tiene los mismos rasgos que el genotipo homocigoto AA ), sin embargo se puede estimar a partir de la frecuencia del rasgo recesivo en la población, ya que esta es la misma que la del genotipo homocigoto aa. es decir, las frecuencias alélicas individuales se pueden estimar: q = √ f (aa), p = 1 − q, y de ellas se puede derivar la frecuencia del genotipo portador: f (Aa) = 2 pq.
Esta fórmula se basa en una serie de suposiciones y una estimación precisa de la frecuencia del rasgo recesivo. En general, cualquier situación del mundo real se desviará de estos supuestos hasta cierto punto, introduciendo las imprecisiones correspondientes en la estimación. Si el rasgo recesivo es raro, será difícil estimar su frecuencia con precisión, ya que se necesitará un tamaño de muestra muy grande.
Dominante versus ventajoso
La propiedad de "dominante" a veces se confunde con el concepto de ventajoso y la propiedad de "recesivo" a veces se confunde con el concepto de perjudicial, pero los fenómenos son distintos. La dominancia describe el fenotipo de los heterocigotos con respecto a los fenotipos de los homocigotos y sin tener en cuenta el grado en que los diferentes fenotipos pueden ser beneficiosos o perjudiciales. Dado que muchos alelos de enfermedades genéticas son recesivos y debido a que la palabra dominancia tiene una connotación positiva, a menudo se asume que el fenotipo dominante es superior con respecto a la aptitud. Sin embargo, esto no está asegurado; como se analiza a continuación, mientras que la mayoría de los alelos de enfermedades genéticas son perjudiciales y recesivos, no todas las enfermedades genéticas son recesivas.
Sin embargo, esta confusión ha sido generalizada a lo largo de la historia de la genética y persiste hasta el día de hoy. Abordar esta confusión fue una de las principales motivaciones para la publicación del principio de Hardy-Weinberg.
Mecanismos moleculares
Mendel desconocía la base molecular de la dominancia. Ahora se entiende que un locus genético incluye una larga serie (cientos a miles) de bases o nucleótidos de ácido desoxirribonucleico (ADN) en un punto particular de un cromosoma. El dogma central de la biología molecular establece que " el ADN hace que el ARN haga proteína ", es decir, que el ADN se transcribe para hacer una copia de ARN, y el ARN se traduce para hacer una proteína. En este proceso, diferentes alelos en un locus pueden o no transcribirse y, si se transcribe, pueden traducirse a versiones ligeramente diferentes de la misma proteína (llamadas isoformas). Las proteínas a menudo funcionan como enzimas que catalizan reacciones químicas en la célula, que directa o indirectamente producen fenotipos. Las mutaciones dentro del genoma pueden alterar la actividad catalítica y, por lo tanto, afectar la dominancia.En cualquier organismo diploide, las secuencias de ADN de los dos alelos presentes en cualquier locus génico pueden ser idénticas (homocigotos) o diferentes (heterocigotos). Incluso si el locus del gen es heterocigoto al nivel de la secuencia de ADN, las proteínas producidas por cada alelo pueden ser idénticas. En ausencia de cualquier diferencia entre los productos proteicos, no se puede decir que ningún alelo sea dominante (ver co-dominancia, arriba). Incluso si los dos productos proteicos son ligeramente diferentes (alozimas), es probable que produzcan el mismo fenotipo con respecto a la acción enzimática y, de nuevo, no se puede decir que ningún alelo sea dominante.
Cigosidad
Históricamente, la ley de la distribución independiente de Mendel suponía que los alelos se clasificarían de forma independiente, siendo un alelo "dominante". La cigosidad, el grado de similitud de los alelos de un organismo, puede afectar la dominancia. Dentro de un organismo diploide, estos estarían definidos por las interacciones haplotípicas de los alelos. La haploidía génica puede resultar en un alelo funcional único que produce suficiente proteína para producir un fenotipo idéntico al del homocigoto.. Son posibles tres tipos generales de interacciones de haplotipos:
- Haplosuficiencia. En un diploide, un alelo funcional de un gen haplosuficiente se consideraría dominante, mientras que un alelo no funcional se consideraría recesivo.Por ejemplo, suponga que la cantidad estándar de enzima producida en el homocigoto funcional es del 100 %, y los dos alelos funcionales contribuyen con el 50 % cada uno. El alelo funcional único en el heterocigoto produce el 50% de la cantidad estándar de enzima, que es suficiente para producir el fenotipo estándar. Si el heterocigoto y el homocigoto con alelo funcional tienen fenotipos idénticos, el alelo funcional es dominante sobre el alelo no funcional. Esto ocurre en el locus del gen albino: el heterocigoto produce suficiente enzima para convertir el precursor del pigmento en melanina y el individuo tiene una pigmentación estándar. Por ejemplo, en humanos y otros organismos,
- Haploinsuficiencia incompleta. Con menos frecuencia, la presencia de un solo alelo funcional da un fenotipo que no es normal, pero menos grave que el del homocigoto no funcional. Esto ocurre cuando el alelo funcional no es haplo-suficiente, por lo que los términos haplo-insuficiencia y dominancia incompleta se aplican típicamente a estos casos. La interacción intermedia ocurre cuando el genotipo heterocigoto produce un fenotipo intermedio entre los dos homocigotos. Según a cuál de los dos homocigotos se parezca más el heterocigoto, se dice que un alelo muestra dominancia incompleta sobre el otro. Por ejemplo, en humanos la HbEl locus del gen es responsable de la proteína de cadena beta (HBB), que es una de las dos proteínas de globina que forman el pigmento sanguíneo de la hemoglobina. Muchas personas son homocigóticas para un alelo llamado Hb ; algunas personas portan un alelo alternativo llamado Hb, ya sea como homocigotos o heterocigotos. Las moléculas de hemoglobina de los homocigotos de Hb / Hb experimentan un cambio de forma que distorsiona la morfología de los glóbulos rojos y provoca una forma de anemia grave y potencialmente mortal llamada anemia de células falciformes. Las personas heterocigotas Hb / Hb para este alelo tienen una forma de anemia mucho menos severa llamada rasgo drepanocítico. Porque el fenotipo de enfermedad de Hb / Hbheterocigotos es más similar pero no idéntico al homocigoto Hb / Hb, se dice que el alelo Hb es incompletamente dominante con respecto al alelo Hb.
- Haploinsuficiencia completa.Un solo alelo funcional en el heterocigoto puede producir un producto génico insuficiente para cualquier función del gen, lo que hace que los alelos generalmente no funcionales se vuelvan dominantes. Entonces, el fenotipo se parecerá al de un homocigoto con alelo no funcional en lugar del tipo salvaje. Se diría que el alelo no funcional es dominante con respecto al alelo funcional del fenotipo de tipo salvaje. Esta situación puede ocurrir cuando el alelo no funcional produce una proteína defectuosa que interfiere con la función adecuada de la proteína producida por el alelo estándar. La presencia de la proteína defectuosa "domina" a la proteína estándar, y el fenotipo de enfermedad del heterocigoto se asemeja más al del homocigoto para dos alelos defectuosos. El término "dominante" a menudo se aplica incorrectamente a los alelos defectuosos cuyo fenotipo homocigoto no se ha examinado, pero que causan un fenotipo distinto cuando son heterocigotos con el alelo normal. Este fenómeno se produce en una serie de enfermedades de repetición de trinucleótidos, siendo un ejemplo la enfermedad de Huntington. En la enfermedad de Huntington, se observa una haploinsuficiencia completa en la que una mutación provoca una enfermedad por un efecto dominante de la proteína mutante.Otro ejemplo es el síndrome de Marfan, un trastorno hereditario del tejido conectivo causado por una mutación en el gen de la fibrilina-1 (FBN1). Una copia normal del gen FBN1 se hereda de uno de los padres, mientras que una copia anormal dominante del gen FBN1 la hereda otro padre.
Mutaciones dominantes negativas
Muchas proteínas son normalmente activas en forma de multímero, un agregado de múltiples copias de la misma proteína, también conocida como proteína homomultimérica o proteína homooligomérica. De hecho, la mayoría de las 83 000 enzimas diferentes de 9800 organismos diferentes en la base de datos de enzimas BRENDA representan homooligómeros. Cuando la versión de tipo salvaje de la proteína está presente junto con una versión mutante, se puede formar un multímero mixto. Una mutación que conduce a una proteína mutante que interrumpe la actividad de la proteína de tipo salvaje en el multímero es una mutación negativa dominante.
Una mutación negativa dominante puede surgir en una célula somática humana y proporcionar una ventaja proliferativa a la célula mutante, lo que lleva a su expansión clonal. Por ejemplo, una mutación negativa dominante en un gen necesario para el proceso normal de muerte celular programada (apoptosis) en respuesta al daño del ADN puede hacer que la célula sea resistente a la apoptosis. Esto permitirá la proliferación del clon incluso cuando esté presente un daño excesivo en el ADN. Tales mutaciones negativas dominantes ocurren en el gen supresor de tumores p53. La proteína de tipo salvaje P53 normalmente está presente como un multímero de cuatro proteínas (oligotetrámero). Las mutaciones p53 negativas dominantes ocurren en varios tipos diferentes de cáncer y lesiones precancerosas (por ejemplo, tumores cerebrales, cáncer de mama, lesiones precancerosas orales y cáncer oral).
Las mutaciones negativas dominantes también ocurren en otros genes supresores de tumores. Por ejemplo, se identificaron dos mutaciones de línea germinal negativas dominantes en el gen mutante de Ataxia telangiectasia (ATM) que aumenta la susceptibilidad al cáncer de mama. Las mutaciones negativas dominantes del factor de transcripción C/EBPα pueden causar leucemia mieloide aguda. Las mutaciones negativas dominantes heredadas también pueden aumentar el riesgo de enfermedades distintas del cáncer. Las mutaciones negativas dominantes en el receptor gamma activado por el proliferador de peroxisomas (PPARγ) están asociadas con resistencia severa a la insulina, diabetes mellitus e hipertensión.
También se han descrito mutaciones negativas dominantes en organismos distintos de los humanos. De hecho, el primer estudio que informó sobre una proteína mutante que inhibía la función normal de una proteína de tipo salvaje en un multímero mixto fue con la proteína GP37 de la fibra de la cola del bacteriófago T4. Las mutaciones que producen una proteína truncada en lugar de una proteína mutante de longitud completa parecen tener el efecto negativo dominante más fuerte en los estudios de P53, ATM, C/EBPα y el bacteriófago T4 GP37.
 Los defectos hereditarios en las enzimas generalmente se heredan de manera autosómica porque hay más cromosomas que no son X que cromosomas X, y de manera recesiva porque las enzimas de los genes no afectados generalmente son suficientes para prevenir los síntomas en los portadores. Las excepciones incluyen casos de haploinsuficiencia, donde el gen no afectado no puede compensar al afectado.
Los defectos hereditarios en las enzimas generalmente se heredan de manera autosómica porque hay más cromosomas que no son X que cromosomas X, y de manera recesiva porque las enzimas de los genes no afectados generalmente son suficientes para prevenir los síntomas en los portadores. Las excepciones incluyen casos de haploinsuficiencia, donde el gen no afectado no puede compensar al afectado.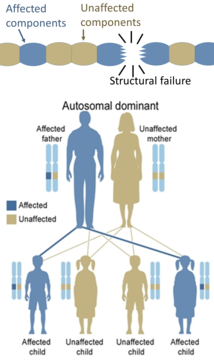 Por otro lado, los defectos hereditarios en las proteínas estructurales (como la osteogénesis imperfecta, el síndrome de Marfan y los síndromes de Ehlers-Danlos) son generalmente autosómicos dominantes, porque basta que algunos componentes sean defectuosos para que toda la estructura sea disfuncional. Este es un proceso negativo dominante, en el que un producto génico mutado afecta negativamente al producto génico no mutado dentro de la misma célula.
Por otro lado, los defectos hereditarios en las proteínas estructurales (como la osteogénesis imperfecta, el síndrome de Marfan y los síndromes de Ehlers-Danlos) son generalmente autosómicos dominantes, porque basta que algunos componentes sean defectuosos para que toda la estructura sea disfuncional. Este es un proceso negativo dominante, en el que un producto génico mutado afecta negativamente al producto génico no mutado dentro de la misma célula.
Enfermedades genéticas dominantes y recesivas en humanos
En los humanos, muchos rasgos genéticos o enfermedades se clasifican simplemente como "dominantes" o "recesivos". Especialmente con las llamadas enfermedades recesivas, que de hecho son un factor de genes recesivos, pero pueden simplificar demasiado la base molecular subyacente y conducir a una mala comprensión de la naturaleza de la dominancia.
Por ejemplo, la enfermedad genética recesiva fenilcetonuria (PKU) resulta de cualquiera de un gran número (>60) de alelos en el locus del gen para la enzima fenilalanina hidroxilasa ( PAH ). Muchos de estos alelos producen poco o nada de PAH, como resultado de lo cual el sustrato fenilalanina (Phe) y sus subproductos metabólicos se acumulan en el sistema nervioso central y pueden causar una discapacidad intelectual grave si no se tratan.
Para ilustrar estos matices, los genotipos y las consecuencias fenotípicas de las interacciones entre tres alelos hipotéticos de PAH se muestran en la siguiente tabla:
| Genotipo | actividad PAH | [ Phe ] concentración | ¿PKU? |
|---|---|---|---|
| Automóvil club británico | 100% | 60 micras | No |
| AB | 30% | 120 micras | No |
| CC | 5% | 200 ~ 300 micras | Hiperfenilalaninemia |
| cama y desayuno | 0,3% | 600 ~ 2400 micras | Sí |
En personas no afectadas homocigóticas para un alelo funcional estándar ( AA ), la actividad de PAH es estándar (100 %) y la concentración de fenilalanina en la sangre [ Phe ] es de aproximadamente 60 μM (= μmol/L). En personas no tratadas homocigóticas para uno de los alelos de PKU ( BB ), la actividad de PAH es cercana a cero, [Phe] de diez a cuarenta veces el estándar, y el individuo manifiesta PKU.
En el heterocigoto AB, la actividad de PAH es solo el 30 % (no el 50 %) del estándar, la [ Phe ] en sangre se eleva al doble y la persona no manifiesta PKU. Por lo tanto, el alelo A es dominante sobre el alelo B con respecto a la PKU, pero el alelo B es incompletamente dominante sobre el alelo A con respecto a su efecto molecular, determinación del nivel de actividad de PAH (0,3 % < 30 % << 100 %). Finalmente, el alelo A es incompletamente dominante al Balelo con respecto a [Phe], como 60 μM < 120 μM << 600 μM. Nótese una vez más que es irrelevante para la cuestión de la dominancia que el alelo recesivo produzca un fenotipo [Phe] más extremo.
Para un tercer alelo C, un homocigoto CC produce una cantidad muy pequeña de enzima PAH, lo que da como resultado un nivel algo elevado de [ Phe ] en la sangre, una condición llamada hiperfenilalaninemia, que no da como resultado una discapacidad intelectual.
Es decir, las relaciones de dominancia de dos alelos cualesquiera pueden variar según el aspecto del fenotipo que se esté considerando. Por lo general, es más útil hablar sobre las consecuencias fenotípicas de las interacciones alélicas involucradas en cualquier genotipo, en lugar de tratar de forzarlas a categorías dominantes y recesivas.
Contenido relacionado
Yodotironina desyodasa
Extinción masiva del Holoceno
Protista