
Descendencia común
Ascendencia común es un concepto en biología evolutiva aplicable cuando una especie es el ancestro de dos o más especies más adelante en el tiempo. Todos los seres vivos son, de hecho, descendientes de un ancestro único comúnmente conocido como el último ancestro común universal (LUCA) de toda la vida en la Tierra, según la biología evolutiva moderna.
La descendencia común es un efecto de la especiación, en el que múltiples especies derivan de una sola población ancestral. Cuanto más reciente es la población ancestral que tienen en común dos especies, más estrechamente están relacionadas. El ancestro común más reciente de todos los organismos vivos actualmente es el último ancestro universal, que vivió hace unos 3.900 millones de años. Las dos pruebas más antiguas de vida en la Tierra son el grafito biogénico encontrado en rocas metasedimentarias de 3.700 millones de años descubiertas en el oeste de Groenlandia y los fósiles microbianos encontrados en areniscas de 3.480 millones de años descubiertas en el oeste de Australia. Todos los organismos que actualmente viven en la Tierra comparten una herencia genética común, aunque la sugerencia de una transferencia de genes horizontal sustancial durante la evolución temprana ha llevado a cuestionar la monofilia (ancestro único) de la vida. Se han identificado 6.331 grupos de genes comunes a todos los animales vivos; estos pueden haber surgido de un solo ancestro común que vivió hace 650 millones de años en el Precámbrico.
La ascendencia común universal a través de un proceso evolutivo fue propuesta por primera vez por el naturalista británico Charles Darwin en la oración final de su libro de 1859 Sobre el origen de las especies:
Hay grandeza en esta visión de la vida, con sus varios poderes, habiendo sido originalmente transpirado en unas cuantas formas o en una; y que, mientras este planeta ha ido ciclándose según la ley fija de la gravedad, de tan simples formas infinitas de principio más hermosas y maravillosas han sido, y están siendo, evolucionados.
Historia
La idea de que todos los seres vivos (incluidos los que la ciencia considera no vivos) están relacionados es un tema recurrente en muchas cosmovisiones indígenas de todo el mundo. Más tarde, en la década de 1740, el matemático francés Pierre Louis Maupertuis llegó a la idea de que todos los organismos tenían un ancestro común y habían divergido por variación aleatoria y selección natural. En Essai de cosmologie (1750), Maupertuis señaló:
Que no digamos eso, en la combinación fortuita de las producciones de la Naturaleza, ya que sólo esas criaturas podría ser sobrevivir en cuyas organizaciones estaba presente un cierto grado de adaptación, no hay nada extraordinario en el hecho de que tal adaptación se encuentra realmente en todas estas especies que existen ahora? Chance, uno podría decir, resultó un gran número de individuos; una pequeña proporción de estos se organizaron de tal manera que los órganos de los animales pudieran satisfacer sus necesidades. Un número mucho mayor no mostró adaptación ni orden; estos últimos han perecido... Así las especies que vemos hoy son sólo una pequeña parte de todos aquellos que ha producido un destino ciego.
En 1790, el filósofo Immanuel Kant escribió en Kritik der Urteilskraft (Crítica del juicio) que la similitud de las formas animales implica un tipo original común y, por lo tanto, un común padre.
En 1794, el abuelo de Charles Darwin, Erasmus Darwin preguntó:
[¿Debería ser demasiado audaz imaginar, que en la gran longitud del tiempo, desde que la tierra comenzó a existir, tal vez millones de edades antes del comienzo de la historia de la humanidad, sería demasiado audaz imaginar, que todos los animales de sangre caliente han surgido de un filamento viviente, que la gran Primera Causa dotada de animalidad, con el poder de adquirir nuevas partes atendidas con nuevas propensiones, dirigidas por irritaciones, sensaciones, voliciones y asociaciones; y así poseer la facultad de seguir mejorando por su propia actividad inherente, y de entregar esas mejoras por generación a su posteridad, mundo sin fin?
Las opiniones de Charles Darwin sobre la descendencia común, expresadas en Sobre el origen de las especies, eran que era probable que solo hubiera un progenitor para todas las formas de vida:
Por lo tanto, debo inferir de analogía que probablemente todos los seres orgánicos que han vivido en esta tierra han descendido de alguna forma primordial, en la que la vida fue respirada por primera vez.
Pero él precede ese comentario con: 'La analogía me llevaría un paso más allá, es decir, a la creencia de que todos los animales y plantas han descendido de algún prototipo. Pero la analogía puede ser una guía engañosa." Y en la edición posterior, afirma más bien,
"No conocemos todas las posibles gradas transicionales entre los órganos más simples y perfectos; no se puede pretender que conozcamos todos los medios variados de Distribución durante el largo lapso de años, o que sabemos lo imperfecto que es el Registro Geológico. Grave como estas varias dificultades son, en mi juicio no derrocan la teoría del descenso de algunas formas creadas con posterior modificación".
La ascendencia común fue ampliamente aceptada entre la comunidad científica después de la publicación de Darwin. En 1907, Vernon Kellogg comentó que "prácticamente ningún naturalista de posición y logros reconocidos duda de la teoría de la descendencia".
En 2008, el biólogo T. Ryan Gregory señaló que:
No se ha encontrado ninguna observación fiable que contradiga la noción general de ascendencia común. No debe sorprender, pues, que la comunidad científica en general haya aceptado el descenso evolutivo como una realidad histórica desde el tiempo de Darwin y la considere entre los hechos más fiablemente establecidos y fundamentalmente importantes en toda la ciencia.
Evidencia
Bioquímica común
Todas las formas de vida conocidas se basan en la misma organización bioquímica fundamental: información genética codificada en ADN, transcrita en ARN, a través del efecto de enzimas de proteínas y ARN, luego traducida en proteínas por ribosomas (muy similares), con ATP, NADPH y otros como fuentes de energía. El análisis de pequeñas diferencias de secuencia en sustancias ampliamente compartidas, como el citocromo c, respalda aún más la ascendencia común universal. Unas 23 proteínas se encuentran en todos los organismos, sirviendo como enzimas que llevan a cabo funciones centrales como la replicación del ADN. El hecho de que solo exista un conjunto de enzimas de este tipo es una evidencia convincente de un solo ancestro. Se han identificado 6.331 genes comunes a todos los animales vivos; estos pueden haber surgido de un solo ancestro común que vivió hace 650 millones de años en el Precámbrico.
Código genético común
| Aminoácidos | nonpolar | polar | básicos | ácido | ¡Detente! |
| 1a base | 2a base | |||||||
|---|---|---|---|---|---|---|---|---|
| T | C | A | G | |||||
| T | TTT | Phenyl-alanine | TCT | Serine | TAT | Tyrosine | TGT | Cysteine |
| TTC | TCC | TAC | TGC | |||||
| TTA | Leucine | TCA | TAA | Para. | TGA | Para. | ||
| TTG | TCG | TAG | Para. | TG | Tryptophan | |||
| C | CTT | CCT | Proline | CAT | Histidina | CGT | Arginine | |
| CTC | CCC | CAC | CGC | |||||
| CTA | CCA | CAA | Glutamina | CGA | ||||
| CTG | CCG | CAG | CG | |||||
| A | ATT | Isoleucine | ACTO | Threonine | AAT | Asparagine | AGT | Serine |
| ATC | ACC | AAC | AGC | |||||
| ATA | ACA | AAA | Lysine | AGA | Arginine | |||
| ATG | Metionina | ACG | AAG | AGG | ||||
| G | GTT | Valine | GCT | Alanine | GAT | Asparticacid | GGT | Glycine |
| GTC | GCC | GAC | GGC | |||||
| GTA | GCA | GAA | Glutamicacid | GGA | ||||
| GTG | GCG | GAG | GG | |||||
El código genético (la "tabla de traducción" según la cual la información del ADN se traduce en aminoácidos y, por lo tanto, en proteínas) es casi idéntico para todas las formas de vida conocidas, desde bacterias y arqueas hasta animales y plantas. Los biólogos generalmente consideran que la universalidad de este código es una evidencia definitiva a favor de la descendencia común universal.
La forma en que los codones (tripletes de ADN) se asignan a los aminoácidos parece estar muy optimizada. Richard Egel argumenta que, en particular, las cadenas laterales hidrofóbicas (no polares) están bien organizadas, lo que sugiere que permitieron a los primeros organismos crear péptidos con regiones repelentes al agua capaces de soportar las reacciones esenciales de intercambio de electrones (redox) para la transferencia de energía.
Similitudes selectivamente neutrales
Las similitudes que no tienen relevancia adaptativa no pueden explicarse mediante la evolución convergente y, por lo tanto, brindan un apoyo convincente para la descendencia común universal. Tal evidencia proviene de dos áreas: secuencias de aminoácidos y secuencias de ADN. Las proteínas con la misma estructura tridimensional no necesitan tener secuencias de aminoácidos idénticas; cualquier similitud irrelevante entre las secuencias es evidencia de descendencia común. En ciertos casos, hay varios codones (tripletes de ADN) que codifican redundantemente para el mismo aminoácido. Dado que muchas especies usan el mismo codón en el mismo lugar para especificar un aminoácido que puede representarse por más de un codón, eso es evidencia de que comparten un ancestro común reciente. Si las secuencias de aminoácidos hubieran venido de ancestros diferentes, habrían sido codificadas por cualquiera de los codones redundantes, y dado que los aminoácidos correctos ya habrían estado en su lugar, la selección natural no habría provocado ningún cambio en los codones, por mucho que lo hicieran. el tiempo estaba disponible. La deriva genética podría cambiar los codones, pero sería muy poco probable que todos los codones redundantes en una secuencia completa coincidieran exactamente en varios linajes. De manera similar, las secuencias de nucleótidos compartidas, especialmente cuando son aparentemente neutrales, como el posicionamiento de los intrones y los pseudogenes, proporcionan una fuerte evidencia de ascendencia común.
Otras similitudes
Los biólogos a menudo señalan la universalidad de muchos aspectos de la vida celular como evidencia de apoyo a la evidencia más convincente mencionada anteriormente. Estas similitudes incluyen el portador de energía trifosfato de adenosina (ATP) y el hecho de que todos los aminoácidos que se encuentran en las proteínas son zurdos. Sin embargo, es posible que estas similitudes se deban a las leyes de la física y la química, en lugar de a través de la descendencia común universal, y por lo tanto dieron como resultado una evolución convergente. Por el contrario, existe evidencia de homología de las subunidades centrales de las ATPasas transmembrana en todos los organismos vivos, especialmente en cómo los elementos giratorios se unen a la membrana. Esto apoya la suposición de un LUCA como un organismo celular, aunque las membranas primordiales pueden haber sido semipermeables y evolucionar más tarde a las membranas de las bacterias modernas, y también en un segundo camino a las de las arqueas modernas.
Árboles filogenéticos
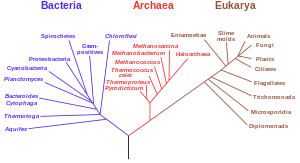
Otra pieza importante de evidencia proviene de árboles filogenéticos detallados (es decir, "árboles genealógicos" de especies) que trazan las divisiones propuestas y los ancestros comunes de todas las especies vivas. En 2010, Douglas L. Theobald publicó un análisis estadístico de los datos genéticos disponibles, asignándolos a árboles filogenéticos, que proporcionó un "fuerte apoyo cuantitativo, mediante una prueba formal, para la unidad de la vida".
Tradicionalmente, estos árboles se han construido utilizando métodos morfológicos, como apariencia, embriología, etc. Recientemente, ha sido posible construir estos árboles utilizando datos moleculares, basados en similitudes y diferencias entre secuencias genéticas y proteicas. Todos estos métodos producen resultados esencialmente similares, aunque la mayor parte de la variación genética no tiene influencia sobre la morfología externa. Que los árboles filogenéticos basados en diferentes tipos de información concuerden entre sí es una fuerte evidencia de una ascendencia común real subyacente.
Posibles objeciones
Análisis filogenético de nubes de intercambio de genes
Theobald señaló que una transferencia genética horizontal sustancial podría haber ocurrido durante la evolución temprana. Hoy en día, las bacterias siguen siendo capaces de intercambiar genes entre linajes relacionados de forma lejana. Esto debilita la suposición básica del análisis filogenético, que la similitud de los genomas implica una ascendencia común, porque un intercambio de genes suficiente permitiría que los linajes compartan gran parte de su genoma, ya sea que compartan o no un antepasado (monofilia). Esto ha llevado a preguntas sobre la ascendencia única de la vida. Sin embargo, los biólogos consideran muy poco probable que protoorganismos completamente no relacionados puedan haber intercambiado genes, ya que sus diferentes mecanismos de codificación habrían resultado solo en sistemas distorsionados en lugar de funcionales. Más tarde, sin embargo, muchos organismos derivados de un solo ancestro podrían haber compartido fácilmente genes que funcionaban todos de la misma manera, y parece que así fue.
Evolución convergente
Si los primeros organismos hubieran sido impulsados por las mismas condiciones ambientales para desarrollar una bioquímica similar de manera convergente, podrían haber adquirido secuencias genéticas similares de manera independiente. La "prueba formal" de Theobald En consecuencia, fue criticado por Takahiro Yonezawa y sus colegas por no incluir la consideración de la convergencia. Argumentaron que la prueba de Theobald era insuficiente para distinguir entre las hipótesis en competencia. Theobald ha defendido su método contra esta afirmación, argumentando que sus pruebas distinguen entre la estructura filogenética y la mera similitud de secuencia. Por lo tanto, argumentó Theobald, sus resultados muestran que "las proteínas reales universalmente conservadas son homólogas".
Mundo ARN
La posibilidad mencionada anteriormente es que todos los organismos vivos pueden descender de un organismo unicelular original con un genoma de ADN, y que esto implica un origen único para la vida. Aunque tal ancestro común universal pudo haber existido, es poco probable que una entidad tan compleja haya surgido espontáneamente de la no vida y, por lo tanto, una célula con un genoma de ADN no puede considerarse razonablemente como el "origen" de la vida. Para comprender el “origen” de la vida, se ha propuesto que la vida celular basada en el ADN descendió de moléculas de ARN autoreplicantes precelulares relativamente simples capaces de someterse a la selección natural (ver el mundo del ARN). Durante el curso de la evolución, este mundo de ARN fue reemplazado por el surgimiento evolutivo del mundo de ADN. Aparentemente, ya no existe un mundo de genomas de ARN autorreplicantes independientes (los virus de ARN dependen de células huésped con genomas de ADN). Debido a que el mundo del ARN aparentemente se ha ido, no está claro cómo la evidencia científica podría influir en la cuestión de si hubo un único "origen" de la vida del que descendió toda la vida.
Contenido relacionado
Trigo escanda
John Eccles (neurofisiólogo)
Alexander Fleming