
Deriva genética
La deriva genética, efecto Wright o desplazamiento genético es el cambio en la frecuencia de una variante genética existente (alelo) en una población debido al azar.
La deriva genética puede hacer que las variantes genéticas desaparezcan por completo y, por lo tanto, reducir la variación genética. También puede hacer que los alelos inicialmente raros se vuelvan mucho más frecuentes e incluso fijos.
Cuando existen pocas copias de un alelo, el efecto de la deriva genética es más notable, y cuando existen muchas copias, el efecto es menos notable. A mediados del siglo XX, se produjeron intensos debates sobre la importancia relativa de la selección natural frente a los procesos neutrales, incluida la deriva genética. Ronald Fisher, quien explicó la selección natural utilizando la genética mendeliana, sostuvo la opinión de que la deriva genética juega un papel menor en la evolución, y esta siguió siendo la opinión dominante durante varias décadas. En 1968, el genetista de poblaciones Motoo Kimura reavivó el debate con su teoría neutral de la evolución molecular, que afirma que la mayoría de los casos en los que un cambio genético se propaga a través de una población (aunque no necesariamente cambios en los fenotipos) son causados por la deriva genética que actúa sobre mutaciones neutrales.En la década de 1990 se propuso la evolución neutral constructiva que busca explicar cómo surgen sistemas complejos a través de transiciones neutrales.
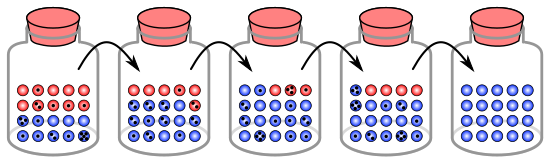
Analogía con canicas en un frasco
El proceso de deriva genética se puede ilustrar usando 20 canicas en un frasco para representar 20 organismos en una población.Considere este frasco de canicas como la población inicial. La mitad de las canicas en el frasco son rojas y la otra mitad azules, y cada color corresponde a un alelo diferente de un gen en la población. En cada nueva generación, los organismos se reproducen al azar. Para representar esta reproducción, selecciona al azar una canica del frasco original y deposita una nueva canica del mismo color en un nuevo frasco. Esta es la "descendencia" de la canica original, lo que significa que la canica original permanece en su frasco. Repita este proceso hasta que haya 20 canicas nuevas en el segundo frasco. El segundo frasco ahora contendrá 20 "descendencia", o canicas de varios colores. A menos que el segundo frasco contenga exactamente 10 canicas rojas y 10 canicas azules, se ha producido un cambio aleatorio en las frecuencias alélicas.
Si este proceso se repite varias veces, el número de canicas rojas y azules recogidas en cada generación fluctúa. A veces, un frasco tiene más canicas rojas que su frasco "principal" y, a veces, más azules. Esta fluctuación es análoga a la deriva genética: un cambio en la frecuencia de alelos de la población que resulta de una variación aleatoria en la distribución de alelos de una generación a la siguiente.
En cualquier generación, no se pueden elegir canicas de un color en particular, lo que significa que no tienen descendencia. En este ejemplo, si no se seleccionan canicas rojas, el frasco que representa la nueva generación contiene solo descendencia azul. Si esto sucede, el alelo rojo se ha perdido permanentemente en la población, mientras que el alelo azul restante se ha vuelto fijo:; todas las generaciones futuras son enteramente azules. En poblaciones pequeñas, la fijación puede ocurrir en unas pocas generaciones.
 En esta simulación, cada punto negro en una canica significa que ha sido elegida para copiar (reproducir) una vez. la fijación en el "alelo" azul ocurre dentro de cinco generaciones.
En esta simulación, cada punto negro en una canica significa que ha sido elegida para copiar (reproducir) una vez. la fijación en el "alelo" azul ocurre dentro de cinco generaciones.
Probabilidad y frecuencia alélica
Los mecanismos de la deriva genética se pueden ilustrar con un ejemplo simplificado. Considere una colonia muy grande de bacterias aisladas en una gota de solución. Las bacterias son genéticamente idénticas excepto por un solo gen con dos alelos etiquetados como A y B, que son alelos neutrales, lo que significa que no afectan la capacidad de la bacteria para sobrevivir y reproducirse; todas las bacterias de esta colonia tienen la misma probabilidad de sobrevivir y reproducirse. Suponga que la mitad de las bacterias tienen el alelo A y la otra mitad tiene el alelo B. Por lo tanto, A y B tienen cada uno una frecuencia alélica de 1/2.
La gota de solución luego se encoge hasta que solo tiene suficiente comida para sostener cuatro bacterias. Todas las demás bacterias mueren sin reproducirse. Entre los cuatro que sobreviven, existen 16 combinaciones posibles para los alelos A y B: (AAAA), (BAAA), (ABAA), (BBAA),(AABA), (BABA), (ABBA), (BBBA),(AAAB), (BAAB), (ABAB), (BBAB),(AABB), (BABB), (ABBB), (BBBB).
Dado que todas las bacterias en la solución original tienen la misma probabilidad de sobrevivir cuando la solución se encoge, los cuatro sobrevivientes son una muestra aleatoria de la colonia original. La probabilidad de que cada uno de los cuatro sobrevivientes tenga un alelo dado es 1/2, por lo que la probabilidad de que ocurra cualquier combinación de alelos en particular cuando la solución se encoge es{displaystyle {frac {1}{2}}cdot {frac {1}{2}}cdot {frac {1}{2}}cdot {frac {1}{2}}= { fracción {1}{16}}.}

(El tamaño de la población original es tan grande que el muestreo ocurre efectivamente con reemplazo). En otras palabras, cada una de las 16 posibles combinaciones de alelos tiene la misma probabilidad de ocurrir, con una probabilidad de 1/16.
Contando las combinaciones con el mismo número de A y B da la siguiente tabla:
| UN | B | combinaciones | Probabilidad |
| 4 | 0 | 1 | 1/16 |
| 3 | 1 | 4 | 4/16 |
| 2 | 2 | 6 | 6/16 |
| 1 | 3 | 4 | 4/16 |
| 0 | 4 | 1 | 1/16 |
Como se muestra en la tabla, el número total de combinaciones que tienen el mismo número de alelos A que de alelos B es seis, y la probabilidad de esta combinación es 6/16. El número total de otras combinaciones es diez, por lo que la probabilidad de un número desigual de alelos A y B es 10/16. Así, aunque la colonia original comenzó con un número igual de A y Balelos, muy posiblemente, el número de alelos en la población restante de cuatro miembros no será igual. La situación de números iguales es en realidad menos probable que la de números desiguales. En este último caso, la deriva genética se ha producido porque las frecuencias alélicas de la población han cambiado debido al muestreo aleatorio. En este ejemplo, la población se contrajo a solo cuatro sobrevivientes aleatorios, un fenómeno conocido como cuello de botella de población.
Las probabilidades del número de copias del alelo A (o B) que sobreviven (dadas en la última columna de la tabla anterior) se pueden calcular directamente a partir de la distribución binomial, donde la probabilidad de "éxito" (probabilidad de que un alelo dado esté presente) es 1/2 (es decir, la probabilidad de que haya k copias de los alelos A (o B) en la combinación) viene dada por:{displaystyle {n elegir k}left({frac {1}{2}}right)^{k}left(1-{frac {1}{2}}right)^{nk }={n elegir k}left({frac {1}{2}}right)^{n}!}

donde n=4 es el número de bacterias supervivientes.
Modelos matemáticos
Los modelos matemáticos de deriva genética se pueden diseñar utilizando procesos de ramificación o una ecuación de difusión que describa los cambios en la frecuencia de alelos en una población idealizada.
Modelo de Wright-Fisher
Considere un gen con dos alelos, A o B. En diploidía, las poblaciones que consisten en N individuos tienen 2 N copias de cada gen. Un individuo puede tener dos copias del mismo alelo o dos alelos diferentes. A un alelo se le asigna la frecuencia p y al otro q. El modelo de Wright-Fisher (llamado así por Sewall Wright y Ronald Fisher) supone que las generaciones no se superponen (por ejemplo, las plantas anuales tienen exactamente una generación por año) y que cada copia del gen que se encuentra en la nueva generación se extrae de forma independiente al azar. de todas las copias del gen en la vieja generación. La fórmula para calcular la probabilidad de obtener kcopias de un alelo que tenía frecuencia p en la última generación es entonces{displaystyle {frac {(2N)!}{k!(2N-k)!}}p^{k}q^{2N-k}}

donde el símbolo " ! " representa la función factorial. Esta expresión también se puede formular usando el coeficiente binomial,{displaystyle {2N elegir k}p^{k}q^{2N-k}}

Modelo morán
El modelo de Moran asume generaciones superpuestas. En cada paso de tiempo, se elige un individuo para reproducirse y un individuo para morir. Entonces, en cada paso de tiempo, el número de copias de un alelo dado puede aumentar en uno, disminuir en uno o permanecer igual. Esto significa que la matriz de transición es tridiagonal, lo que significa que las soluciones matemáticas son más fáciles para el modelo de Moran que para el modelo de Wright-Fisher. Por otro lado, las simulaciones por computadora suelen ser más fáciles de realizar con el modelo de Wright-Fisher, porque es necesario calcular menos pasos de tiempo. En el modelo de Moran, se necesitan N pasos de tiempo para pasar por una generación, donde N es el tamaño efectivo de la población. En el modelo de Wright-Fisher, solo se necesita uno.
En la práctica, los modelos de Moran y Wright-Fisher dan resultados cualitativamente similares, pero la deriva genética corre el doble de rápido en el modelo de Moran.
Otros modelos de deriva
Si la varianza en el número de descendientes es mucho mayor que la dada por la distribución binomial asumida por el modelo de Wright-Fisher, entonces dada la misma velocidad general de la deriva genética (la varianza del tamaño efectivo de la población), la deriva genética es una fuerza menos poderosa. en comparación con la selección. Incluso para la misma varianza, si los momentos más altos de la distribución del número de descendientes superan a los de la distribución binomial, entonces nuevamente la fuerza de la deriva genética se debilita sustancialmente.
Efectos aleatorios distintos del error de muestreo
Los cambios aleatorios en las frecuencias alélicas también pueden ser causados por efectos distintos al error de muestreo, por ejemplo, cambios aleatorios en la presión de selección.
Una importante fuente alternativa de estocasticidad, quizás más importante que la deriva genética, es el borrador genético. El draft genético es el efecto sobre un locus por selección en loci enlazados. Las propiedades matemáticas del borrador genético son diferentes de las de la deriva genética. La dirección del cambio aleatorio en la frecuencia alélica se autocorrelaciona entre generaciones.
Deriva y fijación
El principio de Hardy-Weinberg establece que dentro de poblaciones suficientemente grandes, las frecuencias alélicas permanecen constantes de una generación a la siguiente a menos que el equilibrio se altere por migración, mutaciones genéticas o selección.
Sin embargo, en poblaciones finitas, no se obtienen nuevos alelos del muestreo aleatorio de alelos pasados a la siguiente generación, pero el muestreo puede causar la desaparición de un alelo existente. Debido a que el muestreo aleatorio puede eliminar, pero no reemplazar, un alelo, y debido a que las disminuciones o aumentos aleatorios en la frecuencia de alelos influyen en las distribuciones de alelos esperadas para la próxima generación, la deriva genética conduce a una población hacia la uniformidad genética con el tiempo. Cuando un alelo alcanza una frecuencia de 1 (100%) se dice que está "fijo" en la población y cuando un alelo alcanza una frecuencia de 0 (0%) se pierde. Las poblaciones más pequeñas logran la fijación más rápido, mientras que en el límite de una población infinita no se logra la fijación. Una vez que un alelo se fija, la deriva genética se detiene, y la frecuencia del alelo no puede cambiar a menos que se introduzca un nuevo alelo en la población a través de una mutación o flujo de genes. Por lo tanto, aunque la deriva genética es un proceso aleatorio y sin dirección, actúa para eliminar la variación genética a lo largo del tiempo.
Tasa de cambio de frecuencia alélica debido a la deriva
Suponiendo que la deriva genética es la única fuerza evolutiva que actúa sobre un alelo, después de t generaciones en muchas poblaciones replicadas, comenzando con frecuencias alélicas de p y q, la variación en la frecuencia alélica entre esas poblaciones es{displaystyle V_{t}approx pqleft(1-exp left(-{frac {t}{2N_{e}}}right)right)}

Tiempo de fijación o pérdida
Suponiendo que la deriva genética es la única fuerza evolutiva que actúa sobre un alelo, en un momento dado, la probabilidad de que un alelo finalmente quede fijo en la población es simplemente su frecuencia en la población en ese momento. Por ejemplo, si la frecuencia p para el alelo A es del 75 % y la frecuencia q para el alelo B es del 25 %, entonces, dado un tiempo ilimitado, la probabilidad de que A finalmente se quede fijo en la población es del 75 % y la probabilidad de que B se quede fijo es 25%.
El número esperado de generaciones para que ocurra la fijación es proporcional al tamaño de la población, de modo que se predice que la fijación ocurrirá mucho más rápidamente en poblaciones más pequeñas. Normalmente se utiliza el tamaño efectivo de la población, que es menor que la población total, para determinar estas probabilidades. La población efectiva (N e) tiene en cuenta factores como el nivel de consanguinidad, la etapa del ciclo de vida en la que la población es más pequeña y el hecho de que algunos genes neutros están ligados genéticamente a otros que están en proceso de selección. El tamaño efectivo de la población puede no ser el mismo para todos los genes de la misma población.
Una fórmula prospectiva utilizada para aproximar el tiempo esperado antes de que un alelo neutral se fije a través de la deriva genética, según el modelo de Wright-Fisher, es{displaystyle {bar {T}}_{text{fijo}}={frac {-4N_{e}(1-p)ln(1-p)}{p}}}

donde T es el número de generaciones, N e es el tamaño efectivo de la población y p es la frecuencia inicial para el alelo dado. El resultado es el número de generaciones que se espera que pasen antes de que ocurra la fijación para un alelo dado en una población con tamaño (N e) y frecuencia alélica (p) dados.
El tiempo esperado para que el alelo neutral se pierda a través de la deriva genética se puede calcular como{displaystyle {bar {T}}_{text{perdido}}={frac {-4N_{e}p}{1-p}}ln p.}

Cuando una mutación aparece solo una vez en una población lo suficientemente grande como para que la frecuencia inicial sea insignificante, las fórmulas se pueden simplificar a{displaystyle {bar {T}}_{text{fijo}}=4N_{e}}

para el número medio de generaciones esperadas antes de la fijación de una mutación neutra, y{displaystyle {bar {T}}_{text{perdido}}=2left({frac {N_{e}}{N}}right)ln(2N)}

para el número medio de generaciones esperadas antes de la pérdida de una mutación neutra.
Tiempo hasta la pérdida tanto con la deriva como con la mutación
Las fórmulas anteriores se aplican a un alelo que ya está presente en una población y que no está sujeto a mutación ni selección natural. Si un alelo se pierde por mutación con mucha más frecuencia de lo que se gana por mutación, entonces la mutación, así como la deriva, pueden influir en el tiempo hasta la pérdida. Si el alelo propenso a la pérdida mutacional comienza fijo en la población y se pierde por mutación a una tasa m por replicación, entonces el tiempo esperado en generaciones hasta su pérdida en una población haploide está dado por{displaystyle {bar {T}}_{text{lost}}approx {begin{cases}{dfrac {1}{m}},{text{ if }}mN_{e}ll 1\[8pt]{dfrac {ln {(mN_{e})}+gamma }{m}}{text{ if }}mN_{e}gg 1end{casos}}}
![{displaystyle {bar {T}}_{text{lost}}approx {begin{cases}{dfrac {1}{m}},{text{ if }}mN_{e}ll 1\[8pt]{dfrac {ln {(mN_{e})}+gamma }{m}}{text{ if }}mN_{e}gg 1end{casos}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/0dca9b7dc746a7b2a8d8a63770ac53782d1639e3)
donde{ estilo de visualización gamma}
Contra la selección natural
En las poblaciones naturales, la deriva genética y la selección natural no actúan aisladamente; ambos fenómenos están siempre en juego, junto con la mutación y la migración. La evolución neutral es el producto tanto de la mutación como de la deriva, no solo de la deriva. De manera similar, incluso cuando la selección supera la deriva genética, solo puede actuar sobre la variación que proporciona la mutación.
Mientras que la selección natural tiene una dirección, guiando la evolución hacia adaptaciones hereditarias al entorno actual, la deriva genética no tiene dirección y está guiada únicamente por las matemáticas del azar. Como resultado, la deriva actúa sobre las frecuencias genotípicas dentro de una población sin tener en cuenta sus efectos fenotípicos. Por el contrario, la selección favorece la propagación de alelos cuyos efectos fenotípicos aumentan la supervivencia y/o reproducción de sus portadores, disminuye las frecuencias de alelos que causan rasgos desfavorables e ignora los que son neutrales.
La ley de los grandes números predice que cuando el número absoluto de copias del alelo es pequeño (p. ej., en poblaciones pequeñas), la magnitud de la deriva en las frecuencias alélicas por generación es mayor. La magnitud de la deriva es lo suficientemente grande como para superar la selección en cualquier frecuencia alélica cuando el coeficiente de selección es menor que 1 dividido por el tamaño efectivo de la población. Por lo tanto, se considera que la evolución no adaptativa resultante del producto de la mutación y la deriva genética es un mecanismo consecuente de cambio evolutivo principalmente dentro de poblaciones pequeñas y aisladas. Las matemáticas de la deriva genética dependen del tamaño efectivo de la población, pero no está claro cómo se relaciona esto con el número real de individuos en una población.El enlace genético con otros genes que están bajo selección puede reducir el tamaño efectivo de la población experimentado por un alelo neutral. A mayor tasa de recombinación, el ligamiento disminuye y con ello este efecto local sobre el tamaño efectivo de la población. Este efecto es visible en los datos moleculares como una correlación entre la tasa de recombinación local y la diversidad genética, y una correlación negativa entre la densidad de genes y la diversidad en las regiones de ADN no codificantes. La estocasticidad asociada con la vinculación a otros genes que están bajo selección no es lo mismo que el error de muestreo y, a veces, se conoce como borrador genético para distinguirlo de la deriva genética.
La baja frecuencia de alelos hace que los alelos sean más vulnerables a ser eliminados por casualidad, anulando incluso la influencia de la selección natural. Por ejemplo, mientras que las mutaciones desventajosas suelen eliminarse rápidamente en grandes poblaciones, las nuevas mutaciones ventajosas son casi tan vulnerables a la pérdida por deriva genética como las mutaciones neutras. Hasta que la frecuencia del alelo para la mutación ventajosa alcance un cierto umbral, la deriva genética no tendrá efecto.
Cuello de botella de población
Un cuello de botella poblacional es cuando una población se contrae a un tamaño significativamente más pequeño durante un corto período de tiempo debido a algún evento ambiental aleatorio. En un verdadero cuello de botella poblacional, las probabilidades de supervivencia de cualquier miembro de la población son puramente aleatorias y no mejoran por ninguna ventaja genética inherente en particular. El cuello de botella puede resultar en cambios radicales en las frecuencias alélicas, completamente independientes de la selección.
El impacto de un cuello de botella demográfico puede mantenerse, incluso cuando el cuello de botella es causado por un evento único, como una catástrofe natural. Un ejemplo interesante de un cuello de botella que causa una distribución genética inusual es la proporción relativamente alta de individuos con daltonismo total de células de bastón (acromatopsia) en el atolón de Pingelap en Micronesia. Después de un cuello de botella, aumenta la endogamia. Esto aumenta el daño causado por las mutaciones deletéreas recesivas, en un proceso conocido como depresión endogámica. Las peores de estas mutaciones se seleccionan en contra, lo que lleva a la pérdida de otros alelos que están genéticamente vinculados a ellas, en un proceso de selección de fondo.Para mutaciones recesivas dañinas, esta selección puede potenciarse como consecuencia del cuello de botella, debido a la depuración genética. Esto conduce a una mayor pérdida de diversidad genética. Además, una reducción sostenida en el tamaño de la población aumenta la probabilidad de que se produzcan más fluctuaciones de alelos en las generaciones venideras.
La variación genética de una población puede reducirse en gran medida por un cuello de botella, e incluso las adaptaciones beneficiosas pueden eliminarse de forma permanente. La pérdida de variación deja a la población sobreviviente vulnerable a nuevas presiones de selección, como enfermedades, cambios climáticos o cambios en la fuente de alimento disponible, porque la adaptación en respuesta a los cambios ambientales requiere suficiente variación genética en la población para que tenga lugar la selección natural.
Ha habido muchos casos conocidos de cuello de botella en la población en el pasado reciente. Antes de la llegada de los europeos, las praderas de América del Norte eran el hábitat de millones de gallinas de las praderas. Solo en Illinois, su número se desplomó de alrededor de 100 millones de aves en 1900 a alrededor de 50 aves en la década de 1990. La disminución de la población fue el resultado de la caza y la destrucción del hábitat, pero una consecuencia ha sido la pérdida de la mayor parte de la diversidad genética de la especie. El análisis de ADN que compara aves de mediados de siglo con aves de la década de 1990 documenta una fuerte disminución en la variación genética en las últimas décadas. Actualmente, el pollo de la pradera mayor está experimentando un bajo éxito reproductivo.
Sin embargo, la pérdida genética causada por el cuello de botella y la deriva genética puede aumentar la aptitud, como en Ehrlichia.
La caza excesiva también provocó un grave cuello de botella en la población del elefante marino del norte en el siglo XIX. Su disminución resultante en la variación genética se puede deducir comparándola con la del elefante marino del sur, que no fue cazado tan agresivamente.
Efecto fundador
El efecto fundador es un caso especial de cuello de botella poblacional, que ocurre cuando un pequeño grupo en una población se separa de la población original y forma una nueva. Se espera que la muestra aleatoria de alelos en la nueva colonia recién formada tergiverse enormemente la población original al menos en algunos aspectos. Incluso es posible que el número de alelos de algunos genes en la población original sea mayor que el número de copias de genes en los fundadores, lo que hace imposible una representación completa. Cuando una colonia recién formada es pequeña, sus fundadores pueden afectar fuertemente la composición genética de la población en el futuro.
Un ejemplo bien documentado se encuentra en la migración Amish a Pensilvania en 1744. Dos miembros de la nueva colonia compartían el alelo recesivo del síndrome de Ellis-Van Creveld. Los miembros de la colonia y sus descendientes tienden a ser aislados religiosos y permanecen relativamente insulares. Como resultado de muchas generaciones de endogamia, el síndrome de Ellis-Van Creveld ahora es mucho más frecuente entre los Amish que en la población general.
La diferencia en las frecuencias de los genes entre la población original y la colonia también puede provocar que los dos grupos se diferencien significativamente en el transcurso de muchas generaciones. A medida que aumenta la diferencia, o distancia genética, las dos poblaciones separadas pueden volverse distintas, tanto genética como fenéticamente, aunque no solo la deriva genética sino también la selección natural, el flujo de genes y la mutación contribuyen a esta divergencia. Este potencial de cambios relativamente rápidos en la frecuencia de los genes de la colonia llevó a la mayoría de los científicos a considerar el efecto fundador (y, por extensión, la deriva genética) como una fuerza impulsora significativa en la evolución de nuevas especies. Sewall Wright fue el primero en atribuir este significado a la deriva aleatoria y a las poblaciones pequeñas y recién aisladas con su teoría de la especiación del equilibrio cambiante.Siguiendo a Wright, Ernst Mayr creó muchos modelos persuasivos para mostrar que la disminución de la variación genética y el pequeño tamaño de la población después del efecto fundador eran de importancia crítica para el desarrollo de nuevas especies. Sin embargo, hoy en día hay mucho menos apoyo para este punto de vista ya que la hipótesis ha sido probada repetidamente a través de investigaciones experimentales y los resultados han sido equívocos en el mejor de los casos.
Historia
El papel del azar en la evolución fue descrito por primera vez por Arend L. Hagedoorn y AC Hagedoorn-Vorstheuvel La Brand en 1921. Destacaron que la supervivencia aleatoria juega un papel clave en la pérdida de variación de las poblaciones. Fisher (1922) respondió a esto con el primer tratamiento matemático, aunque marginalmente incorrecto, del 'efecto Hagedoorn'. En particular, esperaba que muchas poblaciones naturales fueran demasiado grandes (N ~ 10,000) para que los efectos de la deriva fueran sustanciales y pensó que la deriva tendría un efecto insignificante en el proceso evolutivo. El tratamiento matemático corregido y el término "deriva genética" fueron acuñados más tarde por un fundador de la genética de poblaciones, Sewall Wright. Su primer uso del término "deriva" fue en 1929,aunque en ese momento lo estaba usando en el sentido de un proceso dirigido de cambio, o selección natural. La deriva aleatoria por medio del error de muestreo llegó a conocerse como el "efecto Sewall-Wright", aunque nunca se sintió del todo cómodo al ver que se le dio su nombre. Wright se refirió a todos los cambios en la frecuencia de alelos como "deriva constante" (p. ej., selección) o "deriva aleatoria" (p. ej., error de muestreo). "Deriva" pasó a ser adoptado como un término técnico en el sentido estocástico exclusivamente. Hoy en día suele definirse de forma aún más estrecha, en términos de error de muestreo, aunque esta definición estrecha no es universal. Wright escribió que la "restricción de la 'deriva aleatoria' o incluso la 'deriva' a un solo componente, los efectos de los accidentes de muestreo,Sewall Wright consideró el proceso de deriva genética aleatoria mediante un error de muestreo equivalente al de la consanguinidad, pero trabajos posteriores han demostrado que son distintos.
En los primeros días de la síntesis evolutiva moderna, los científicos estaban comenzando a combinar la nueva ciencia de la genética de poblaciones con la teoría de la selección natural de Charles Darwin. Dentro de este marco, Wright se centró en los efectos de la consanguinidad en poblaciones pequeñas relativamente aisladas. Introdujo el concepto de un paisaje adaptativo en el que fenómenos como el cruzamiento y la deriva genética en pequeñas poblaciones podrían alejarlas de los picos adaptativos, lo que a su vez permite que la selección natural las empuje hacia nuevos picos adaptativos. Wright pensó que las poblaciones más pequeñas eran más adecuadas para la selección natural porque "la consanguinidad era lo suficientemente intensa como para crear nuevos sistemas de interacción a través de la deriva aleatoria, pero no lo suficientemente intensa como para causar la fijación aleatoria no adaptativa de genes".
Las opiniones de Wright sobre el papel de la deriva genética en el esquema evolutivo fueron controvertidas casi desde el principio. Uno de los críticos más vociferantes e influyentes fue su colega Ronald Fisher. Fisher admitió que la deriva genética desempeñó algún papel en la evolución, pero insignificante. Fisher ha sido acusado de malinterpretar los puntos de vista de Wright porque en sus críticas, Fisher parecía argumentar que Wright había rechazado la selección casi por completo. Para Fisher, ver el proceso de evolución como una progresión larga, constante y adaptativa era la única forma de explicar la complejidad cada vez mayor a partir de formas más simples. Pero los debates han continuado entre los "gradualistas" y aquellos que se inclinan más hacia el modelo de evolución de Wright, donde la selección y la deriva juntas juegan un papel importante.
En 1968, Motoo Kimura reavivó el debate con su teoría neutral de la evolución molecular, que afirma que la mayoría de los cambios genéticos son causados por la deriva genética que actúa sobre mutaciones neutrales.
El papel de la deriva genética por medio del error de muestreo en la evolución ha sido criticado por John H. Gillespie y William B. Provine, quienes argumentan que la selección en sitios vinculados es una fuerza estocástica más importante.
Contenido relacionado
Simbiogénesis
Lucio del norte
Apareamiento