
Cristalino (anatomía de los vertebrados)
El lente, o cristalino, es una estructura biconvexa transparente que se encuentra en la mayoría de los ojos de los vertebrados terrestres. Junto con la córnea, los humores acuoso y vítreo refracta la luz, enfocándola sobre la retina. En muchos animales terrestres, la forma de la lente se puede alterar, cambiando efectivamente la distancia focal del ojo, lo que les permite enfocar objetos a varias distancias. Este ajuste de la lente se conoce como acomodación (ver también a continuación). En muchos vertebrados totalmente acuáticos, como los peces, se utilizan otros métodos de acomodación, como cambiar la posición del cristalino en relación con la retina, en lugar de cambiar la forma del cristalino. La acomodación es análoga al enfoque de una cámara fotográfica cambiando sus lentes. En los vertebrados terrestres, el cristalino es más plano en su lado anterior que en su lado posterior, mientras que en los peces, el cristalino suele ser casi esférico.
La acomodación en humanos está bien estudiada y permite medios artificiales para complementar nuestro enfoque, como anteojos para corregir la vista a medida que envejecemos. El poder de refracción de una lente humana más joven en su entorno natural es de aproximadamente 18 dioptrías, aproximadamente un tercio del poder total del ojo de unas 60 dioptrías. A los 25 años, la capacidad de la lente para alterar la trayectoria de la luz se ha reducido a 10 dioptrías y la acomodación sigue disminuyendo con la edad.
Estructura
Posición en el ojo
El cristalino está ubicado hacia la parte frontal del ojo de los vertebrados, llamado segmento anterior, que incluye la córnea y el iris colocados frente al cristalino. El cristalino se mantiene en su lugar gracias a los ligamentos suspensorios (Zonule de Zinn), que unen el cristalino en su ecuador con el resto del ojo a través del cuerpo ciliar. Detrás del cristalino se encuentra el cuerpo vítreo gelatinoso que ayuda a mantener el cristalino en su lugar. En la parte frontal del cristalino se encuentra el humor acuoso líquido que baña el cristalino con nutrientes y otras cosas. Los lentes de los vertebrados terrestres suelen tener una forma elipsoide biconvexa. La superficie frontal es menos curva que la trasera. En un adulto humano, el cristalino suele tener unos 10 mm de diámetro y 4 mm de grosor, aunque cambia de forma con la acomodación y el tamaño debido al crecimiento a lo largo de la vida de una persona.
Anatomía




El cristalino tiene tres partes principales: la cápsula del cristalino, el epitelio del cristalino y las fibras del cristalino. La cápsula del cristalino es una membrana basal relativamente gruesa que forma la capa más externa del cristalino. Dentro de la cápsula, fibras mucho más delgadas del cristalino forman la mayor parte del cristalino. Las células del epitelio del cristalino forman una capa delgada entre la cápsula del cristalino y la capa más externa de fibras del cristalino en la parte delantera del cristalino, pero no en la parte trasera. El cristalino en sí mismo carece de nervios, vasos sanguíneos o tejido conectivo. Los anatomistas a menudo se refieren a las posiciones de las estructuras en el cristalino describiéndolo como un globo terráqueo. La parte delantera y trasera del cristalino se denominan "polos" anterior y posterior, al igual que los polos norte y sur. El "ecuador" es el borde exterior del cristalino, a menudo oculto por el iris y es el área de mayor diferenciación celular. Como el ecuador generalmente no está en el camino de la luz del ojo, las estructuras involucradas en la actividad metabólica evitan la dispersión de la luz que, de otro modo, afectaría la visión.
Cápsula del cristalino


La cápsula del cristalino es una membrana basal suave y transparente que rodea completamente el cristalino. La cápsula es elástica y su principal componente estructural es el colágeno. Se presume que es sintetizado por el epitelio del cristalino y sus componentes principales en orden de abundancia son heparán sulfato proteoglicano (glicosaminoglicanos sulfatados (GAG)), entactina, colágeno tipo IV, laminina. La cápsula es muy elástica y permite que el cristalino adopte una forma más esférica cuando se reduce la tensión de los ligamentos suspensorios. La cápsula humana varía de 2 a 28 micrómetros de espesor, siendo más gruesa cerca del ecuador (región periecuatorial) y generalmente más delgada cerca del polo posterior. La foto de un microscopio electrónico muestra un área de la cápsula cerca del ecuador donde se une uno de los miles de ligamentos suspensorios.

La unión debe ser lo suficientemente fuerte como para evitar que el ligamento se separe de la cápsula del cristalino. Las fuerzas se generan al sostener la lente en su lugar y se suman al enfocar. La cápsula anterior y posterior es más delgada.
Epitelio del cristalino
El epitelio del cristalino es una sola capa de células en la parte frontal del cristalino entre la cápsula del cristalino y las fibras del cristalino. Al proporcionar nutrientes a las fibras del cristalino y eliminar los desechos, las células del epitelio regulan el mantenimiento de la homeostasis del cristalino. A medida que los iones, los nutrientes y el líquido ingresan al cristalino desde el humor acuoso, las bombas Na+/K+-ATPasa en las células epiteliales del cristalino bombean iones fuera del cristalino para mantener la concentración y el volumen osmóticos apropiados del cristalino, y las células del epitelio del cristalino ubicadas ecuatorialmente contribuyen más a esta corriente La actividad de las Na+/K+-ATPasas mantiene el agua y la corriente fluyendo a través de la lente desde los polos y saliendo por las regiones ecuatoriales.
Las células del epitelio del cristalino también se dividen en nuevas fibras del cristalino en el ecuador del cristalino. El cristalino deposita fibras desde que se forma por primera vez en el embrión hasta la muerte.
Fibras de lentes
Las fibras del cristalino forman la mayor parte del cristalino. Son células largas, delgadas y transparentes, empaquetadas firmemente, con diámetros típicamente de 4 a 7 micrómetros y longitudes de hasta 12 mm en humanos. Las fibras del cristalino se extienden a lo largo desde el polo posterior hasta el anterior y, cuando se cortan horizontalmente, se disponen en capas concéntricas como las capas de una cebolla. Si se corta a lo largo del ecuador, aparece como un panal. La mitad aproximada de cada fibra se encuentra alrededor del ecuador. Estas capas apretadas de fibras de lentes se denominan láminas. Los citoplasmas de la fibra del cristalino están unidos entre sí a través de uniones comunicantes, puentes intercelulares e interdigitaciones de las células que se asemejan a "bola y cavidad" formularios
La lente se divide en regiones dependiendo de la edad de las fibras de la lente de una capa particular. Moviéndose hacia afuera desde la capa central más antigua, la lente se divide en un núcleo embrionario, el núcleo fetal, el núcleo adulto, la corteza interna y externa. Las nuevas fibras del cristalino, generadas a partir del epitelio del cristalino, se añaden a la corteza exterior. Las fibras maduras del cristalino no tienen orgánulos ni núcleos.
Fusión celular, vacíos y vacuolas


Con el advenimiento de otras formas de observar las estructuras celulares del cristalino mientras aún estaban en el animal vivo, se hizo evidente que las regiones de las células de fibra, al menos en la parte anterior del cristalino, contienen grandes vacíos y vacuolas. Se especula que estos están involucrados en los sistemas de transporte de lentes que unen la superficie de la lente con regiones más profundas. Las estructuras de aspecto muy similar también indican fusión celular en el cristalino. La fusión celular se muestra mediante microinyección para formar un sincitio estratificado en cultivos de lentes completos.
Desarrollo

El desarrollo del cristalino de los vertebrados comienza cuando el embrión humano mide unos 4 mm de largo. La imagen adjunta muestra el proceso en un embrión de pollo más fácil de estudiar. A diferencia del resto del ojo, que se deriva principalmente de las capas internas del embrión, el cristalino se deriva de la piel que rodea al embrión. La primera etapa de la formación del cristalino tiene lugar cuando una esfera de células formada por la gemación de las capas internas del embrión se acerca a la piel externa del embrión. La esfera de células induce a la piel exterior cercana a comenzar a transformarse en la placoda del cristalino. La placoda del cristalino es la primera etapa de transformación de un parche de piel en el cristalino. En esta etapa temprana, la placoda del cristalino es una sola capa de células.
A medida que avanza el desarrollo, la placoda del cristalino comienza a profundizarse y a inclinarse hacia adentro. A medida que la placoda continúa profundizándose, la abertura hacia la superficie del ectodermo se contrae y las células del cristalino brotan de la piel del embrión para formar una esfera de células conocida como "vesícula del cristalino". Cuando el embrión mide unos 10 mm de largo, la vesícula del cristalino se ha separado completamente de la piel del embrión.
Luego, el embrión envía señales desde la retina en desarrollo, lo que induce a las células más cercanas al extremo posterior de la vesícula del cristalino a alargarse hacia el extremo anterior de la vesícula. Estas señales también inducen la síntesis de proteínas llamadas cristalinas. Como sugiere el nombre, los cristalinos pueden formar una gelatina transparente altamente refractiva. Estas células que se alargan eventualmente llenan el centro de la vesícula con células, que son largas y delgadas como un mechón de cabello, llamadas fibras. Estas fibras primarias se convierten en el núcleo del cristalino maduro. Las células epiteliales que no forman fibras más cercanas al frente del cristalino dan origen al epitelio del cristalino.

Las fibras adicionales se derivan de las células epiteliales del cristalino ubicadas en el ecuador del cristalino. Estas células se alargan hacia delante y hacia atrás envolviendo fibras ya depositadas. Las nuevas fibras deben ser más largas para cubrir las fibras anteriores, pero a medida que la lente crece, los extremos de las fibras más nuevas ya no llegan tan lejos hacia la parte delantera y trasera de la lente. Las fibras del cristalino que no llegan a los polos forman costuras estrechas e interdigitadas con las fibras vecinas. Estas costuras, que son menos cristalinas que la mayor parte del cristalino, son más visibles y se denominan "suturas". Los patrones de sutura se vuelven más complejos a medida que se añaden más capas de fibras del cristalino a la parte exterior del cristalino.
El cristalino continúa creciendo después del nacimiento, y las nuevas fibras secundarias se agregan como capas externas. Las nuevas fibras del cristalino se generan a partir de las células ecuatoriales del epitelio del cristalino, en una región denominada "zona germinativa" y "región del arco". Las células epiteliales del cristalino se alargan, pierden contacto con la cápsula y el epitelio en la parte posterior y frontal del cristalino, sintetizan cristalina y finalmente pierden sus núcleos (enuclean) a medida que se convierten en fibras maduras del cristalino. En los seres humanos, a medida que el cristalino crece depositando más fibras hasta la edad adulta temprana, el cristalino adquiere una forma más elipsoide. Después de los 20 años, el cristalino se vuelve más redondo y el iris es muy importante para este desarrollo.
Varias proteínas controlan el desarrollo embrionario del cristalino, aunque PAX6 se considera el gen regulador principal de este órgano. Otros efectores del desarrollo adecuado de la lente incluyen los componentes de señalización Wnt BCL9 y Pygo2. Todo el proceso de diferenciación de las células epiteliales en células fibrosas llenas de cristalino sin orgánulos ocurre dentro de los límites de la cápsula del cristalino. Las células más viejas no se pueden desprender y, en cambio, se internalizan hacia el centro de la lente. Este proceso da como resultado un registro completo en capas temporales del proceso de diferenciación desde el inicio en la superficie de la lente hasta el final en el centro de la lente. Por lo tanto, la lente es valiosa para los científicos que estudian el proceso de diferenciación celular.
Variaciones en la estructura del cristalino

En muchos vertebrados acuáticos, el cristalino es considerablemente más grueso, casi esférico, lo que resulta en una mayor refracción de la luz. Esta diferencia ayuda a compensar el menor ángulo de refracción entre la córnea del ojo y el ambiente acuoso, ya que tienen índices de refracción más similares que la córnea y el aire. Las células de fibra de los peces son generalmente considerablemente más delgadas que las de los vertebrados terrestres y parece que las proteínas cristalinas se transportan a las células libres de orgánulos en el exterior del cristalino a las células internas a través de muchas capas de células. Algunos vertebrados necesitan ver bien tanto por encima como por debajo del agua a veces. Un ejemplo son las aves buceadoras que tienen la capacidad de cambiar el enfoque entre 50 y 80 dioptrías. En comparación con los animales adaptados para un solo entorno, las aves buceadoras tienen una estructura de lente y córnea algo alterada con mecanismos de enfoque para permitir ambos entornos. Incluso entre los animales terrestres, el cristalino de los primates, como los humanos, es inusualmente plano, lo que explica de alguna manera por qué nuestra visión, a diferencia de las aves buceadoras, es particularmente borrosa bajo el agua.
Función
Enfocar


En los seres humanos, el mecanismo de enfoque de Helmholtz ampliamente citado, también llamado acomodación, a menudo se denomina "modelo". La prueba experimental directa de cualquier modelo de lente es necesariamente difícil ya que la lente de los vertebrados es transparente y solo funciona bien en los animales vivos. Al considerar todos los vertebrados, los aspectos de todos los modelos pueden desempeñar diferentes roles en el enfoque de la lente.
La lente que cambia de forma de muchos vertebrados terrestres
Fuerzas externas
Young propuso el modelo de una lente humana que cambia de forma en una conferencia el 27 de noviembre de 1800. Otros, como Helmholtz y Huxley, refinaron el modelo a mediados de 1800 y explicaron cómo se contrae el músculo ciliar redondeando la lente para enfocar de cerca. y este modelo fue popularizado por Helmholtz en 1909. El modelo puede resumirse así. Normalmente, el cristalino se mantiene bajo tensión porque sus ligamentos de suspensión se tensan por la presión del globo ocular. A una distancia focal corta, el músculo ciliar se contrae y alivia parte de la tensión en los ligamentos, lo que permite que el cristalino se redondee un poco elásticamente, lo que aumenta el poder de refracción. Cambiar el enfoque a un objeto a una distancia mayor requiere una lente menos curva y más delgada. Esto se logra relajando algunos de los músculos ciliares del esfínter. Si bien no se hace referencia a esto, presumiblemente permite que la presión en el globo ocular lo expanda nuevamente hacia afuera, tirando con más fuerza de la lente, haciéndola menos curva y más delgada, lo que aumenta la distancia focal. Hay un problema con el modelo de Helmholtz en el sentido de que, a pesar de que se probaron modelos matemáticos, ninguno se ha acercado lo suficiente a trabajar usando solo los mecanismos de Helmholtz.

Schachar ha propuesto un modelo para vertebrados terrestres que no fue bien recibido. La teoría permite que el modelado matemático refleje con mayor precisión la forma en que se enfoca la lente y, al mismo tiempo, tiene en cuenta las complejidades de los ligamentos suspensorios y la presencia de músculos radiales y circulares en el cuerpo ciliar. En este modelo, los ligamentos pueden tirar en diversos grados del cristalino en el ecuador utilizando los músculos radiales, mientras que los ligamentos desplazados desde el ecuador hacia adelante y hacia atrás se relajan en diversos grados al contraer los músculos circulares. Estas múltiples acciones que actúan sobre la lente elástica le permiten cambiar la forma de la lente en la parte delantera de forma más sutil. No solo cambiando el enfoque, sino también corrigiendo las aberraciones de la lente que de otro modo podrían resultar del cambio de forma mientras se ajusta mejor al modelo matemático.
La "catenaria" El modelo de enfoque del cristalino propuesto por Coleman exige menos tensión en los ligamentos que suspenden el cristalino. En lugar de que el cristalino en su conjunto se estire más delgado para la visión de lejos y se le permita relajarse para enfocar de cerca, la contracción de los músculos ciliares circulares da como resultado que el cristalino tenga menos presión hidrostática contra su frente. El frente de la lente puede entonces reformar su forma entre los ligamentos suspensorios de manera similar a como una cadena floja que cuelga entre dos polos podría cambiar su curva cuando los polos se acercan. Este modelo requiere solo un movimiento fluido del frente de la lente en lugar de tratar de cambiar la forma de la lente como un todo.
Fuerzas internas


Cuando Thomas Young propuso el cambio de la forma del cristalino humano como mecanismo para la acomodación focal en 1801, pensó que el cristalino podría ser un músculo capaz de contraerse. Este tipo de modelo se denomina acomodación intracapsular ya que se basa en la actividad dentro del cristalino. En una conferencia del Nobel de 1911, Allvar Gullstrand habló sobre "Cómo descubrí el mecanismo intracapsular de acomodación". y este aspecto del enfoque de la lente continúa siendo investigado. Young pasó tiempo buscando los nervios que podrían estimular la contracción del cristalino sin éxito. Desde entonces ha quedado claro que el cristalino no es un simple músculo estimulado por un nervio, por lo que el modelo de Helmholtz de 1909 tuvo prioridad. Los investigadores anteriores al siglo XX no tuvieron el beneficio de muchos descubrimientos y técnicas posteriores. Las proteínas de membrana, como las acuaporinas, que permiten que el agua entre y salga de las células, son las proteínas de membrana más abundantes en el cristalino. También prevalecen las conexinas que permiten el acoplamiento eléctrico de las células. La microscopía electrónica y la microscopía inmunofluorescente muestran que las células de fibra son muy variables en estructura y composición. La resonancia magnética confirma una capa en el cristalino que puede permitir diferentes planes de refracción en su interior. El índice de refracción del cristalino humano varía desde aproximadamente 1,406 en las capas centrales hasta 1,386 en las capas menos densas del cristalino. Este gradiente de índice mejora la potencia óptica de la lente. A medida que se aprende más sobre la estructura del cristalino de los mamíferos a partir de fotografías de Scheimpflug in situ, resonancias magnéticas e investigaciones fisiológicas, se hace evidente que el cristalino en sí mismo no responde de manera completamente pasiva al músculo ciliar circundante, pero puede cambiar su estado general. índice de refracción a través de mecanismos que involucran la dinámica del agua en la lente aún por aclarar. La micrografía adjunta muestra fibras arrugadas de un cristalino de oveja relajado después de retirarlo del animal, lo que indica un acortamiento de las fibras del cristalino durante la acomodación del foco cercano. Los cambios relacionados con la edad en el cristalino humano también pueden estar relacionados con cambios en la dinámica del agua en el cristalino.
Lentes de aves, reptiles, anfibios, peces y otras
(feminine)
En reptiles y aves, el cuerpo ciliar que soporta el cristalino a través de los ligamentos suspensorios también toca el cristalino con una serie de almohadillas en su superficie interna. Estas almohadillas comprimen y liberan la lente para modificar su forma mientras enfoca objetos a diferentes distancias; los ligamentos suspensorios suelen realizar esta función en los mamíferos. En la visión de los peces y los anfibios, el cristalino tiene una forma fija y, en cambio, el enfoque se logra moviendo el cristalino hacia adelante o hacia atrás dentro del ojo usando un músculo llamado retractor lentus.
En los peces cartilaginosos, los ligamentos suspensorios se reemplazan por una membrana, que incluye un pequeño músculo en la parte inferior del cristalino. Este músculo empuja la lente hacia adelante desde su posición relajada cuando enfoca objetos cercanos. En los teleósteos, por el contrario, un músculo se proyecta desde una estructura vascular en el piso del ojo, llamado proceso falciforme, y sirve para tirar de la lente hacia atrás desde la posición relajada para enfocar objetos distantes. Mientras que los anfibios mueven el cristalino hacia adelante, al igual que los peces cartilaginosos, los músculos involucrados no son similares en ninguno de los dos tipos de animales. En las ranas, hay dos músculos, uno arriba y otro debajo del cristalino, mientras que otros anfibios solo tienen el músculo inferior.
En los vertebrados más simples, las lampreas y los mixinos, el cristalino no está unido en absoluto a la superficie exterior del globo ocular. No hay humor acuoso en estos peces, y el cuerpo vítreo simplemente presiona el cristalino contra la superficie de la córnea. Para enfocar sus ojos, una lamprea aplana la córnea usando músculos fuera del ojo y empuja la lente hacia atrás.
Aunque no es un vertebrado, aquí se hace una breve mención de la evolución convergente de los ojos de los vertebrados y los moluscos. El ojo de los moluscos más complejo es el ojo de los cefalópodos, que tiene una estructura y una función superficialmente similares a las de un ojo de vertebrado, incluida la acomodación, aunque difiere en aspectos básicos, como tener una lente de dos partes y no tener córnea. Los requisitos fundamentales de la óptica deben ser cumplidos por todos los ojos con lentes utilizando los tejidos a su disposición para que superficialmente todos los ojos tiendan a verse similares. Es la forma en que se cumplen los requisitos ópticos utilizando diferentes tipos de células y mecanismos estructurales que varía entre los animales.
Cristalinas y transparencia

Los cristalinos son proteínas solubles en agua que componen más del 90 % de la proteína dentro del cristalino. Los tres tipos principales de cristales que se encuentran en el ojo humano son los cristales α, β y γ. Los cristalinos tienden a formar agregados solubles de alto peso molecular que se empaquetan estrechamente en las fibras de la lente, lo que aumenta el índice de refracción de la lente y mantiene su transparencia. Los cristalinos β y γ se encuentran principalmente en el cristalino, mientras que las subunidades de cristalino α se han aislado de otras partes del ojo y del cuerpo. Las proteínas α-cristalinas pertenecen a una superfamilia más grande de proteínas chaperonas moleculares, por lo que se cree que las proteínas cristalinas se reclutaron evolutivamente a partir de proteínas chaperonas con fines ópticos. Las funciones de chaperona de la α-cristalina también pueden ayudar a mantener las proteínas del cristalino, que deben durar toda la vida de un ser humano.
Otro factor importante para mantener la transparencia del cristalino es la ausencia de orgánulos que dispersen la luz, como el núcleo, el retículo endoplásmico y las mitocondrias dentro de las fibras maduras del cristalino. Las fibras del cristalino también tienen un citoesqueleto muy extenso que mantiene la forma precisa y el empaquetamiento de las fibras del cristalino; las interrupciones/mutaciones en ciertos elementos del citoesqueleto pueden conducir a la pérdida de transparencia.
La lente bloquea la mayor parte de la luz ultravioleta en el rango de longitud de onda de 300 a 400 nm; las longitudes de onda más cortas son bloqueadas por la córnea. El pigmento responsable de bloquear la luz es el glucósido de 3-hidroxiquinurenina, un producto del catabolismo del triptófano en el epitelio del cristalino. La luz ultravioleta de alta intensidad puede dañar la retina y, por lo tanto, se fabrican lentes intraoculares artificiales para bloquear también la luz ultravioleta. Las personas que carecen de un cristalino (una condición conocida como afaquia) perciben la luz ultravioleta como azul blanquecino o violeta blanquecino.
Alimentación
El cristalino es metabólicamente activo y requiere nutrición para mantener su crecimiento y transparencia. Sin embargo, en comparación con otros tejidos del ojo, el cristalino tiene una demanda de energía considerablemente menor.
A las nueve semanas del desarrollo humano, el cristalino está rodeado y alimentado por una red de vasos, la túnica vasculosa lentis, que se deriva de la arteria hialoidea. A partir del cuarto mes de desarrollo, la arteria hialoidea y su vasculatura relacionada comienzan a atrofiarse y desaparecen por completo al nacer. En el ojo posnatal, el canal de Cloquet marca la ubicación anterior de la arteria hialoidea.

Después de la regresión de la arteria hialoidea, el cristalino recibe toda su nutrición del humor acuoso. Los nutrientes se difunden y los desechos se difunden a través de un flujo constante de líquido desde los polos anterior/posterior del cristalino y fuera de las regiones ecuatoriales, una dinámica que es mantenida por el Na+/K +-bombas de ATPasa ubicadas en las células del epitelio del cristalino situadas en posición ecuatorial. La interacción de estas bombas con canales de agua en células llamadas acuaporinas, moléculas de menos de 100 daltons de tamaño entre las células a través de uniones comunicantes, y transportadores/reguladores que usan calcio (canales TRPV) da como resultado un flujo de nutrientes a través de la lente.
La glucosa es la principal fuente de energía del cristalino. Como las fibras maduras del cristalino no tienen mitocondrias, aproximadamente el 80 % de la glucosa se metaboliza a través del metabolismo anaeróbico. La fracción restante de glucosa se desvía principalmente por la vía de las pentosas fosfato. La falta de respiración aeróbica hace que el cristalino consuma muy poco oxígeno.
Importancia clínica
- Las cataratas son opacidades de la lente. Aunque algunos son pequeños y no requieren ningún tratamiento, otros pueden ser lo suficientemente grandes para bloquear la luz y obstruir la visión. Las cataratas suelen desarrollarse a medida que la lente de envejecimiento se vuelve cada vez más opaca, pero las cataratas también pueden formar congénitamente o después de la lesión en la lente. La esclerosis nuclear es un tipo de catarata relacionada con la edad. La diabetes es otro factor de riesgo para la catarata. La cirugía de catarata implica la eliminación de la lente e inserción de una lente intraocular artificial.
- Presbyopia es la pérdida de alojamiento relacionada con la edad, que está marcada por la incapacidad del ojo para centrarse en objetos cercanos. El mecanismo exacto sigue siendo desconocido, pero los cambios relacionados con la edad en la dureza, la forma y el tamaño de la lente se han relacionado con la condición.
- Ectopia lentis es el desplazamiento de la lente desde su posición normal.
- Aphakia es la ausencia de la lente del ojo. Aphakia puede ser el resultado de cirugía o lesión, o puede ser congénita.
Imágenes adicionales

Escaneo de resonancia magnética de los ojos humanos.
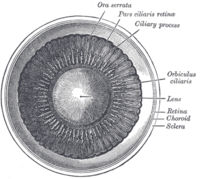
Interior de cámara anterior del ojo.

La lente cristalina, endurecida y dividida.

Sección a través del margen de la lente, mostrando la transición del epitelio a las fibras de lente conocidas como la región del arco.
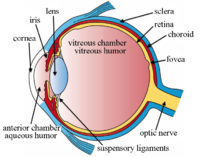
Las estructuras del ojo etiquetadas

Otra vista del ojo y las estructuras del ojo etiquetadas
![This svg file was configured so that the rays, diaphragm and crystalline lens are easily modified[69]](https://upload.wikimedia.org/wikipedia/commons/thumb/5/5e/Editable_ray_diagram_of_eye_v0.svg/200px-Editable_ray_diagram_of_eye_v0.svg.png)
Este archivo svg fue configurado para que los rayos, diafragma y lente cristalino sean fácilmente modificados





![This svg file was configured so that the rays, diaphragm and crystalline lens are easily modified[69]](https://en.wikipedia.org/wiki/File:Editable_ray_diagram_of_eye_v0.svg)
Contenido relacionado
Gadiformes
Mancha norteña
Monoamina oxidasa