
CRISPR
CRISPR () (un acrónimo de cs lustered rregularmente iinterspaced srepeticiones palindrómicas cortas) es una familia de secuencias de ADN que se encuentran en los genomas de organismos procarióticos como bacterias y arqueas. Estas secuencias se derivan de fragmentos de ADN de bacteriófagos que habían infectado previamente al procariota. Se utilizan para detectar y destruir el ADN de bacteriófagos similares durante infecciones posteriores. Por lo tanto, estas secuencias desempeñan un papel clave en el sistema de defensa antiviral (es decir, antifagos) de los procariotas y proporcionan una forma de inmunidad adquirida. CRISPR se encuentra en aproximadamente el 50% de los genomas bacterianos secuenciados y en casi el 90% de las arqueas secuenciadas.

Cas9 (o "proteína 9 asociada a CRISPR") es una enzima que utiliza secuencias CRISPR como guía para reconocer y abrir hebras específicas de ADN que son complementarias a la secuencia CRISPR. Las enzimas Cas9 junto con las secuencias CRISPR forman la base de una tecnología conocida como CRISPR-Cas9 que puede usarse para editar genes dentro de los organismos. Este proceso de edición tiene una amplia variedad de aplicaciones que incluyen investigación biológica básica, desarrollo de productos biotecnológicos y tratamiento de enfermedades. El desarrollo de la técnica de edición del genoma CRISPR-Cas9 fue reconocido con el Premio Nobel de Química en 2020 otorgado a Emmanuelle Charpentier y Jennifer Doudna.
Historia
Secuencias repetidas
El descubrimiento de repeticiones de ADN agrupadas se produjo de forma independiente en tres partes del mundo. La primera descripción de lo que más tarde se llamaría CRISPR la realizó el investigador de la Universidad de Osaka Yoshizumi Ishino y sus colegas en 1987. Clonaron accidentalmente parte de una secuencia CRISPR junto con el "iap" gen (conversión de isoenzimas de fosfatasa alcalina) del genoma de Escherichia coli que era su objetivo. La organización de las repeticiones fue inusual. Las secuencias repetidas normalmente se organizan de forma consecutiva, sin intercalar secuencias diferentes. No conocían la función de las repeticiones agrupadas interrumpidas.
En 1993, investigadores de Mycobacterium tuberculosis in the Netherlands published two articles about a cluster of interrupted direct repeats (DR) in that bacterium. Reconocieron la diversidad de las secuencias que intervinieron en las repeticiones directas entre diferentes cepas de M. tuberculosis y usó esta propiedad para diseñar un método de escritura que fue nombrado Spoligotyping, que todavía está en uso, hoy.
Francisco Mojica de la Universidad de Alicante en España estudió las repeticiones observadas en los organismos arqueales de las especies Haloferax y Haloarcula y su función. El supervisor de Mojica supuso en ese momento que las repeticiones agrupadas tenían un papel en la segregación correcta del ADN replicado en células hijas durante la división celular porque los plásmidos y cromosomas con matrices de repeticiones idénticas no podían coexistir en Haloferax volcanii. También se observó por primera vez la transcripción de las repeticiones interrumpidas; esta fue la primera caracterización completa de CRISPR. Hacia el año 2000, Mojica realizó un estudio de la literatura científica y uno de sus alumnos realizó una búsqueda en genomas publicados con un programa ideado por él mismo. Identificaron repeticiones interrumpidas en 20 especies de microbios como pertenecientes a la misma familia. Debido a que esas secuencias estaban espaciadas, Mojica inicialmente las llamó "repeticiones cortas regularmente espaciadas" (SRSR). En 2001, Mojica y Ruud Jansen, que buscaban repeticiones interrumpidas adicionales, propusieron el acrónimo CRISPR (Clustered Regularly Interspaced Short Palindromic Repeticiones) para aliviar la confusión derivada de los numerosos acrónimos utilizados para describir las secuencias en la literatura científica. En 2002, Tang y cols. mostró evidencia de que las regiones repetidas CRISPR del genoma de Archaeoglobus fulgidus se transcribieron en largas moléculas de ARN que posteriormente se procesaron en ARN pequeños de longitud unitaria, además de algunas formas más largas de 2, 3 o más secuencias espaciadoras. repetir unidades.
En 2005, el investigador del yogur Rodolphe Barrangou descubrió que el Streptococcus thermophilus, después de desafíos iterativos con fagos, desarrolla una mayor resistencia a los fagos, y esta resistencia mejorada se debe a la incorporación de secuencias espaciadoras CRISPR adicionales. La empresa alimentaria danesa Danisco, para la que entonces trabajaba Barrangou, desarrolló S resistente a fagos. cepas thermophilus para su uso en la producción de yogur. Posteriormente, Danisco fue comprada por DuPont, que "posee alrededor del 50 por ciento del mercado mundial de cultivo lácteo" y la tecnología se generalizó.
Sistemas asociados a CRISPR
Un avance importante en la comprensión de CRISPR se produjo con la observación de Jansen de que el grupo de repeticiones procariotas estaba acompañado por un conjunto de genes homólogos que conforman los sistemas asociados a CRISPR o genes cas. Inicialmente se reconocieron cuatro genes cas (cas 1–4). Las proteínas Cas mostraron motivos de helicasa y nucleasa, lo que sugiere un papel en la estructura dinámica de los loci CRISPR. En esta publicación se utilizó el acrónimo CRISPR como nombre universal de este patrón. Sin embargo, la función CRISPR seguía siendo enigmática.

En 2005, tres grupos de investigación independientes demostraron que algunos espaciadores CRISPR se derivan de ADN de fagos y ADN extracromosómico, como los plásmidos. En efecto, los espaciadores son fragmentos de ADN obtenidos de virus que previamente intentaron atacar la célula. El origen de los espaciadores fue una señal de que el sistema CRISPR-cas podría tener un papel en la inmunidad adaptativa de las bacterias. Los tres estudios que proponían esta idea fueron rechazados inicialmente por revistas de alto perfil, pero finalmente aparecieron en otras revistas.
La primera publicación que propone un papel de CRISPR-Cas en la inmunidad microbiana, realizada por Mojica y colaboradores de la Universidad de Alicante, predijo un papel de la transcripción de ARN de los espaciadores en el reconocimiento de objetivos en un mecanismo que podría ser análogo al de interferencia del ARN. Sistema utilizado por las células eucariotas. Koonin y sus colegas ampliaron esta hipótesis de interferencia del ARN proponiendo mecanismos de acción para los diferentes subtipos de CRISPR-Cas según la función prevista de sus proteínas.
El trabajo experimental de varios grupos reveló los mecanismos básicos de la inmunidad CRISPR-Cas. En 2007 se publicó la primera evidencia experimental de que CRISPR era un sistema inmunológico adaptativo. Una región CRISPR en Streptococcus thermophilus adquirió espaciadores del ADN de un bacteriófago infectante. Los investigadores manipularon la resistencia de S. thermophilus a diferentes tipos de fagos agregando y eliminando espaciadores cuya secuencia coincidía con la encontrada en los fagos probados. En 2008, Brouns y Van der Oost identificaron un complejo de proteínas Cas (llamado Cascade) que en E. coli cortó el precursor de ARN CRISPR dentro de las repeticiones en moléculas de ARN maduras que contienen espaciadores llamadas ARN CRISPR (ARNcr), que permanecieron unidas al complejo proteico. Además, se descubrió que se necesitaban Cascade, crRNA y una helicasa/nucleasa (Cas3) para proporcionar a un huésped bacteriano inmunidad contra la infección por un virus de ADN. Al diseñar un CRISPR antivirus, demostraron que dos orientaciones del crRNA (sentido/antisentido) proporcionaban inmunidad, lo que indica que las guías del crRNA estaban dirigidas al dsDNA. Ese año Marraffini y Sontheimer confirmaron que una secuencia CRISPR de S. epidermidis se dirigió al ADN y no al ARN para evitar la conjugación. Este hallazgo contradecía el mecanismo propuesto de inmunidad CRISPR-Cas similar a la interferencia del ARN, aunque más tarde se encontró en Pyrococcus furiosus un sistema CRISPR-Cas que se dirige al ARN extraño. Un estudio de 2010 demostró que CRISPR-Cas corta ambas hebras de ADN de fagos y plásmidos en S. thermophilus.
Cas9
Un sistema CRISPR más simple de Streptococcus pyogenes se basa en la proteína Cas9. La endonucleasa Cas9 es un sistema de cuatro componentes que incluye dos moléculas pequeñas: crRNA y ARN CRISPR transactivador (tracrRNA). En 2012, Jennifer Doudna y Emmanuelle Charpentier rediseñaron la endonucleasa Cas9 para convertirla en un sistema de dos componentes más manejable fusionando las dos moléculas de ARN en un "ARN de guía única" que, cuando se combina con Cas9, podría encontrar y cortar el objetivo de ADN especificado por el ARN guía. Esta contribución fue tan significativa que fue reconocida con el Premio Nobel de Química en 2020. Al manipular la secuencia de nucleótidos del ARN guía, el sistema artificial Cas9 podría programarse para apuntar a cualquier secuencia de ADN para su separación. Otro grupo de colaboradores compuesto por Virginijus Šikšnys junto con Gasiūnas, Barrangou y Horvath demostró que Cas9 del S. thermophilus El sistema CRISPR también se puede reprogramar para apuntar a un sitio de su elección cambiando la secuencia de su ARNcr. Estos avances impulsaron los esfuerzos para editar genomas con el sistema CRISPR-Cas9 modificado.
Los grupos liderados por Feng Zhang y George Church publicaron simultáneamente descripciones de la edición del genoma en cultivos de células humanas utilizando CRISPR-Cas9 por primera vez. Desde entonces se ha utilizado en una amplia gama de organismos, incluida la levadura de panadería (Saccharomyces cerevisiae), el patógeno oportunista Candida albicans, el pez cebra (Danio rerio), moscas de la fruta (Drosophila melanogaster), hormigas (Harpegnathos saltator y Ooceraea biroi), mosquitos (Aedes aegypti), nematodos (Caenorhabditis elegans), plantas, ratones (Mus musculus domesticus), monos y embriones humanos.
CRISPR se ha modificado para crear factores de transcripción programables que permiten apuntar y activar o silenciar genes específicos.

Se ha demostrado que el sistema CRISPR-Cas9 realiza ediciones genéticas efectivas en cigotos tripronucleares humanos, descrito por primera vez en un artículo de 2015 de los científicos chinos P. Liang e Y. Xu. El sistema logró escindir con éxito la beta-hemoglobina (HBB) mutante en 28 de 54 embriones. Cuatro de los 28 embriones se recombinaron con éxito utilizando una plantilla de donante proporcionada por los científicos. Los científicos demostraron que durante la recombinación del ADN de la cadena escindida, la secuencia endógena homóloga HBD compite con la plantilla donante exógena. La reparación del ADN en embriones humanos es mucho más complicada y particular que en células madre derivadas.
Cas12a
En 2015, la nucleasa Cas12a (anteriormente conocida como Cpf1) se caracterizó en el sistema CRISPR-Cpf1 de la bacteria Francisella novicida. Su nombre original, a partir de una definición de familia de proteínas TIGRFAM creada en 2012, refleja la prevalencia de su subtipo CRISPR-Cas en los linajes Prevotella y Francisella. Cas12a mostró varias diferencias clave con respecto a Cas9, entre las que se incluyen: provocar un efecto 'escalonado' cortado en ADN bicatenario a diferencia del ADN 'romo' corte producido por Cas9, basándose en un 'T rico' PAM (que proporciona sitios de direccionamiento alternativos a Cas9) y requiere solo un ARN CRISPR (crRNA) para un direccionamiento exitoso. Por el contrario, Cas9 requiere tanto crRNA como un crRNA transactivador (tracrRNA).
Estas diferencias pueden darle a Cas12a algunas ventajas sobre Cas9. Por ejemplo, los pequeños crRNA de Cas12a son ideales para la edición del genoma multiplexado, ya que se pueden empaquetar más de ellos en un vector que los sgRNA de Cas9. Los salientes pegajosos 5' que deja Cas12a también se pueden utilizar para el ensamblaje de ADN, que es mucho más específico para el objetivo que la clonación tradicional con enzimas de restricción. Finalmente, Cas12a escinde el ADN entre 18 y 23 pares de bases aguas abajo del sitio PAM. Esto significa que no hay alteración de la secuencia de reconocimiento después de la reparación, por lo que Cas12a permite múltiples rondas de escisión del ADN. Por el contrario, dado que Cas9 corta sólo 3 pares de bases aguas arriba del sitio PAM, la vía NHEJ produce mutaciones indel que destruyen la secuencia de reconocimiento, evitando así más rondas de corte. En teoría, las rondas repetidas de escisión del ADN deberían generar una mayor oportunidad para que se produzca la edición genómica deseada. Una característica distintiva de Cas12a, en comparación con Cas9, es que después de cortar su objetivo, Cas12a permanece unido al objetivo y luego escinde otras moléculas de ADN ss de forma no discriminatoria. Esta propiedad se llama "escisión colateral" o "trans-escisión" actividad y ha sido aprovechado para el desarrollo de diversas tecnologías de diagnóstico.
Cas13
En 2016, la nucleasa Cas13a (anteriormente conocida como C2c2) de la bacteria Leptotrichia shahii fue caracterizado. Cas13 es una endonucleasa de ARN guiada por ARN, lo que significa que no escinde ADN, sino sólo ARN monocatenario. Cas13 es guiado por su crRNA hacia un objetivo de ssRNA y se une y escinde el objetivo. Al igual que Cas12a, Cas13 permanece unido al objetivo y luego escinde otras moléculas de ssRNA de forma no discriminatoria. Esta propiedad de escisión colateral se ha aprovechado para el desarrollo de diversas tecnologías de diagnóstico.
En 2021, el Dr. Hui Yang caracterizó nuevas variantes en miniatura de la proteína Cas13 (mCas13), Cas13X y Cas13Y. El uso de una pequeña porción de la secuencia del gen N del SARS-CoV-2 como objetivo en la caracterización de mCas13, reveló la sensibilidad y especificidad de mCas13 junto con RT-LAMP para la detección de SARS-CoV-2 en muestras tanto sintéticas como clínicas en comparación con otras pruebas estándar disponibles como RT-qPCR (1 copia/μL).
Estructura del lugar
Repeticiones y espaciadores
La matriz CRISPR se compone de una secuencia líder rica en AT seguida de repeticiones cortas separadas por espaciadores únicos. Las repeticiones CRISPR suelen tener un tamaño de entre 28 y 37 pares de bases (pb), aunque puede haber desde 23 pb hasta 55 pb. Algunos muestran simetría diada, lo que implica la formación de una estructura secundaria como un tallo en bucle ('horquilla') en el ARN, mientras que otros están diseñados para no estar estructurados. El tamaño de los espaciadores en diferentes matrices CRISPR suele ser de 32 a 38 pb (rango de 21 a 72 pb). Pueden aparecer rápidamente nuevos espaciadores como parte de la respuesta inmune a la infección por fagos. Por lo general, hay menos de 50 unidades de la secuencia espaciadora repetida en una matriz CRISPR.
Estructuras de ARN CRISPR
 CRISPR-DR2: Estructura secundaria extraída de la base de datos Rfam. Familia RF01315.
CRISPR-DR2: Estructura secundaria extraída de la base de datos Rfam. Familia RF01315.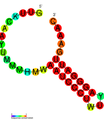 CRISPR-DR5: Estructura secundaria extraída de la base de datos Rfam. Familia RF011318.
CRISPR-DR5: Estructura secundaria extraída de la base de datos Rfam. Familia RF011318. CRISPR-DR6: Estructura secundaria extraída de la base de datos Rfam. Familia RF01319.
CRISPR-DR6: Estructura secundaria extraída de la base de datos Rfam. Familia RF01319. CRISPR-DR8: Estructura secundaria extraída de la base de datos Rfam. Familia RF01321.
CRISPR-DR8: Estructura secundaria extraída de la base de datos Rfam. Familia RF01321. CRISPR-DR9: Estructura secundaria extraída de la base de datos Rfam. Familia RF01322.
CRISPR-DR9: Estructura secundaria extraída de la base de datos Rfam. Familia RF01322.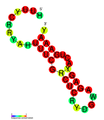 CRISPR-DR19: Estructura secundaria extraída de la base de datos Rfam. Familia RF01332.
CRISPR-DR19: Estructura secundaria extraída de la base de datos Rfam. Familia RF01332.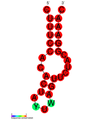 CRISPR-DR41: Estructura secundaria extraída de la base de datos Rfam. Familia RF01350.
CRISPR-DR41: Estructura secundaria extraída de la base de datos Rfam. Familia RF01350. CRISPR-DR52: Estructura secundaria extraída de la base de datos Rfam. Familia RF01365.
CRISPR-DR52: Estructura secundaria extraída de la base de datos Rfam. Familia RF01365.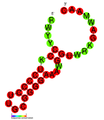 CRISPR-DR57: Estructura secundaria extraída de la base de datos Rfam. Familia RF01370.
CRISPR-DR57: Estructura secundaria extraída de la base de datos Rfam. Familia RF01370. CRISPR-DR65: Estructura secundaria extraída de la base de datos Rfam. Familia RF01378.
CRISPR-DR65: Estructura secundaria extraída de la base de datos Rfam. Familia RF01378.










genes Cas y subtipos CRISPR
Pequeños grupos de genes cas a menudo se ubican junto a las matrices de espaciadores de repetición CRISPR. En conjunto, los 93 genes cas se agrupan en 35 familias según la similitud de secuencia de las proteínas codificadas. 11 de las 35 familias forman el núcleo cas, que incluye las familias de proteínas Cas1 a Cas9. Un locus CRISPR-Cas completo tiene al menos un gen que pertenece al núcleo cas.
Los sistemas CRISPR-Cas se dividen en dos clases. Los sistemas de clase 1 utilizan un complejo de múltiples proteínas Cas para degradar ácidos nucleicos extraños. Los sistemas de clase 2 utilizan una única proteína Cas grande para el mismo propósito. La clase 1 se divide en tipos I, III y IV; La clase 2 se divide en los tipos II, V y VI. Los 6 tipos de sistemas se dividen en 19 subtipos. Cada tipo y la mayoría de los subtipos se caracterizan por un "gen característico" encontrado casi exclusivamente en la categoría. La clasificación también se basa en el complemento de genes cas que están presentes. La mayoría de los sistemas CRISPR-Cas tienen una proteína Cas1. La filogenia de las proteínas Cas1 generalmente concuerda con el sistema de clasificación, pero existen excepciones debido a la combinación de módulos. Muchos organismos contienen múltiples sistemas CRISPR-Cas, lo que sugiere que son compatibles y pueden compartir componentes. La distribución esporádica de los subtipos CRISPR-Cas sugiere que el sistema CRISPR-Cas está sujeto a transferencia horizontal de genes durante la evolución microbiana.
| Clase | Tipo de casco | Subtipo de casco | Proteína de la firma | Función | Referencia |
|---|---|---|---|---|---|
| 1 | I | — | Cas3 | nucleasa de ADN monoplaza (dominio HD) y helicasa dependiente de ATP | |
| I-A | Cas8a, Cas5 | Cas8 es una subunidad del módulo de interferencia que es importante en la selección de ADN invadiendo reconociendo la secuencia PAM. Cas5 es necesario para el procesamiento y estabilidad de crRNAs. | |||
| I-B | Cas8b | ||||
| I-C | Cas8c | ||||
| I-D | Cas10d | contiene un dominio homologoso al dominio de palma de polimeras de ácido nucleico y ciclos de nucleótido | |||
| I-E | Cse1, Cse2 | ||||
| I-F | Csy1, Csy2, Csy3 | Tipo IF-3 han sido implicados en transposones asociados con CRISPR | |||
| I-G | GSU0054 | ||||
| III | — | Cas10 | Homolog of Cas10d and Cse1. Binds CRISPR target RNA y promueve la estabilidad del complejo de interferencia | ||
| III-A | Csm2 | No determinado | |||
| III-B | Cmr5 | No determinado | |||
| III-C | Cas10 o Csx11 | ||||
| III-D | Csx10 | ||||
| III-E | |||||
| III-F | |||||
| IV | — | Csf1 | |||
| IV-A | |||||
| IV-B | |||||
| IV-C | |||||
| 2 | II | — | Cas9 | Nucleases RuvC y HNH juntos producen DSBs, y por separado pueden producir rupturas de una sola pista. Garantiza la adquisición de espaciadores funcionales durante la adaptación. | |
| II-A | Csn2 | Proteína de unión de ADN en forma de anillo. Involved in primed adaptation in Type II CRISPR system. | |||
| II-B | Cas4 | Endonuclease que funciona con cas1 y cas2 para generar secuencias de espacior | |||
| II-C | Caracterizado por la ausencia de Csn2 o Cas4 | ||||
| V | — | Cas12 | Nuclease RuvC. Lacks HNH. | ||
| V-A | Cas12a (Cpf1) | Procesamiento automático de la actividad pre-crRNA para la regulación de genes multiplex | |||
| V-B | Cas12b (C2c1) | ||||
| V-C | Cas12c (C2c3) | ||||
| V-D | Cas12d (CasY) | ||||
| V-E | Cas12e (CasX) | ||||
| V-F | Cas12f (Cas14, C2c10) | ||||
| V-G | Cas12g | ||||
| V-H | Cas12h | ||||
| V-I | Cas12i | ||||
| V-K | Cas12k (C2c5) | El tipo V-K ha estado implicado en transposones asociados con CRISPR. | |||
| V-U | C2c4, C2c8, C2c9 | ||||
| VI | — | Cas13 | RNase guiado por ARN | ||
| VI-A | Cas13a (C2c2) | ||||
| VI-B | Cas13b | ||||
| VI-C | Cas13c | ||||
| VI-D | Cas13d | ||||
| VI-X | Cas13x.1 | Polimerasa RNA dependiente RNA, Inhibición Profiláctica del RNA-virus | |||
| VI-Y |
Mecanismo

(1) La adquisición comienza por el reconocimiento de ADN invadido por Cas1 y Cas2 y el escote de un protoespacial.
(2) El protospacer está ligado a la repetición directa adyacente a la secuencia de líder y
(3) la extensión de hebra única repara el CRISPR y duplica la repetición directa. Las etapas de procesamiento e interferencia del crRNA ocurren de manera diferente en cada uno de los tres sistemas principales del CRISPR.
(4) La transcripción principal de CRISPR es cleaved por genes de cas para producir crRNAs.
(5) En sistemas tipo I Cas6e/Cas6f cleave en la unión de ssRNA y dsRNA formados por los lazos de horquilla en la repetición directa. Los sistemas Tipo II utilizan un ARN transactivador (tracr) para formar dsRNA, que es cleaved por Cas9 y RNaseIII. Los sistemas Tipo III usan un homolog Cas6 que no requiere lazos de horquilla en la repetición directa para el escote.
(6) En los sistemas de tipo II y tipo III se realiza el recortado secundario al final de 5' o 3' para producir crRNAs maduros.
(7) Los crRNAs maduros se asocian con proteínas Cas para formar complejos de interferencia.
(8) En los sistemas tipo I y tipo II, las interacciones entre la secuencia de proteínas y PAM son necesarias para la degradación del ADN invadido. Los sistemas Tipo III no requieren un PAM para la degradación exitosa y en sistemas de base tipo III-A se produce entre el crRNA y el mRNA en lugar del ADN, dirigido por sistemas tipo III-B.





La inmunidad CRISPR-Cas es un proceso natural de bacterias y arqueas. CRISPR-Cas previene la infección, la conjugación y la transformación natural de los bacteriófagos al degradar los ácidos nucleicos extraños que ingresan a la célula.
Adquisición de espaciadores
Cuando un microbio es invadido por un bacteriófago, la primera etapa de la respuesta inmune es capturar el ADN del fago e insertarlo en un locus CRISPR en forma de espaciador. Cas1 y Cas2 se encuentran en ambos tipos de sistemas inmunológicos CRISPR-Cas, lo que indica que están involucrados en la adquisición de espaciadores. Los estudios de mutaciones confirmaron esta hipótesis y demostraron que la eliminación de Cas1 o Cas2 detuvo la adquisición del espaciador, sin afectar la respuesta inmune CRISPR.
Se han caracterizado múltiples proteínas Cas1 y se han resuelto sus estructuras. Las proteínas Cas1 tienen diversas secuencias de aminoácidos. Sin embargo, sus estructuras cristalinas son similares y todas las proteínas Cas1 purificadas son nucleasas/integrasas dependientes de metales que se unen al ADN de manera independiente de la secuencia. Se han caracterizado proteínas Cas2 representativas y poseen actividad endoribonucleasa específica de ARNss (de una sola hebra) o de ADNds (de doble hebra).
En el sistema I-E de E. coli Cas1 y Cas2 forman un complejo en el que un dímero Cas2 une dos dímeros Cas1. En este complejo, Cas2 desempeña una función de andamiaje no enzimático, uniendo fragmentos bicatenarios de ADN invasor, mientras que Cas1 se une a los flancos monocatenarios del ADN y cataliza su integración en matrices CRISPR. Generalmente se agregan nuevos espaciadores al comienzo de CRISPR junto a la secuencia líder creando un registro cronológico de infecciones virales. En E. coli una proteína similar a una histona llamada factor de integración del huésped (IHF), que se une a la secuencia líder, es responsable de la precisión de esta integración. IHF también mejora la eficiencia de la integración en el sistema tipo I-F de Pectobacterium atrosepticum. pero en otros sistemas, es posible que se requieran diferentes factores del huésped
Motivos adyacentes al protoespaciador (PAM)
El análisis bioinformático de regiones de genomas de fagos que fueron extirpados como espaciadores (denominados protoespaciadores) reveló que no fueron seleccionados al azar sino que se encontraron adyacentes a secuencias de ADN cortas (3 a 5 pb) denominadas motivos adyacentes a protoespaciadores (PAM). El análisis de los sistemas CRISPR-Cas mostró que los PAM son importantes para los sistemas tipo I y tipo II, pero no para los sistemas tipo III durante la adquisición. En los sistemas de tipo I y tipo II, los protoespaciadores se escinden en posiciones adyacentes a una secuencia PAM, y el otro extremo del espaciador se corta utilizando un mecanismo de regla, manteniendo así la regularidad del tamaño del espaciador en la matriz CRISPR. La conservación de la secuencia PAM difiere entre los sistemas CRISPR-Cas y parece estar vinculada evolutivamente a Cas1 y la secuencia líder.
Se agregan nuevos espaciadores a una matriz CRISPR de manera direccional, y se producen de manera preferencial, pero no exclusiva, adyacente a la secuencia líder. Análisis del sistema tipo I-E de E. coli demostró que la primera repetición directa adyacente a la secuencia líder se copia, con el espaciador recién adquirido insertado entre la primera y segunda repeticiones directas.
La secuencia PAM parece ser importante durante la inserción del espaciador en sistemas tipo I-E. Esa secuencia contiene un nucleótido final (nt) fuertemente conservado adyacente al primer nt del protoespaciador. Este nt se convierte en la base final en la primera repetición directa. Esto sugiere que la maquinaria de adquisición del espaciador genera salientes monocatenarios en la penúltima posición de la repetición directa y en el PAM durante la inserción del espaciador. Sin embargo, no todos los sistemas CRISPR-Cas parecen compartir este mecanismo ya que los PAM en otros organismos no muestran el mismo nivel de conservación en la posición final. Es probable que en esos sistemas se genere un extremo romo al final de la repetición directa y el protoespaciador durante la adquisición.
Variantes de inserción
El análisis de los CRISPR de Sulfolobus solfataricus reveló mayores complejidades en el modelo canónico de inserción de espaciadores, ya que uno de sus seis loci CRISPR insertó nuevos espaciadores aleatoriamente en toda su matriz CRISPR, en lugar de insertarlos más cerca del líder. secuencia.
Varios CRISPR contienen muchos espaciadores para el mismo fago. El mecanismo que provoca este fenómeno fue descubierto en el sistema tipo I-E de E. coli. Se detectó una mejora significativa en la adquisición de espaciadores cuando los espaciadores ya apuntan al fago, incluso si no coinciden con el protoespaciador. Este 'cebado' requiere que las proteínas Cas involucradas tanto en la adquisición como en la interferencia interactúen entre sí. Los espaciadores recién adquiridos que resultan del mecanismo de cebado siempre se encuentran en el mismo hilo que el espaciador de cebado. Esta observación llevó a la hipótesis de que la maquinaria de adquisición se desliza a lo largo del ADN extraño después del cebado para encontrar un nuevo protoespaciador.
Biogénesis
El ARN CRISPR (ARNcr), que luego guía la nucleasa Cas hacia el objetivo durante el paso de interferencia, debe generarse a partir de la secuencia CRISPR. El crRNA se transcribe inicialmente como parte de una única transcripción larga que abarca gran parte de la matriz CRISPR. Luego, las proteínas Cas escinden esta transcripción para formar crRNA. El mecanismo para producir crRNA difiere entre los sistemas CRISPR-Cas. En los sistemas de tipo I-E y tipo I-F, las proteínas Cas6e y Cas6f, respectivamente, reconocen bucles de tallo creados por el emparejamiento de repeticiones idénticas que flanquean el ARNcr. Estas proteínas Cas escinden el transcrito más largo en el borde de la región emparejada, dejando un único ARNcr junto con un pequeño remanente de la región repetida emparejada.
Los sistemas de tipo III también utilizan Cas6; sin embargo, sus repeticiones no producen bucles de vástago. En cambio, la escisión se produce cuando la transcripción más larga envuelve Cas6 para permitir la escisión justo aguas arriba de la secuencia repetida.
Los sistemas de tipo II carecen del gen Cas6 y en su lugar utilizan RNasaIII para la escisión. Los sistemas funcionales de tipo II codifican un ARN extra pequeño que es complementario a la secuencia repetida, conocido como ARNcr transactivador (ARNtracr). La transcripción del ARNtracr y la transcripción CRISPR primaria da como resultado el emparejamiento de bases y la formación de ARNds en la secuencia repetida, que posteriormente es dirigida por la RNasaIII para producir ARNcr. A diferencia de los otros dos sistemas, el crRNA no contiene el espaciador completo, que en cambio está truncado en un extremo.
Los CrRNA se asocian con las proteínas Cas para formar complejos de ribonucleótidos que reconocen ácidos nucleicos extraños. Los CrRNA no muestran preferencia entre las cadenas codificantes y no codificantes, lo que es indicativo de un sistema de direccionamiento al ADN guiado por ARN. El complejo de tipo I-E (comúnmente conocido como Cascade) requiere cinco proteínas Cas unidas a un solo crRNA.
Interferencia
Durante la etapa de interferencia en los sistemas de tipo I, la secuencia PAM se reconoce en la cadena complementaria del crRNA y se requiere junto con el recocido del crRNA. En los sistemas de tipo I, el emparejamiento de bases correcto entre el crRNA y el protoespaciador señala un cambio conformacional en Cascade que recluta Cas3 para la degradación del ADN.
Los sistemas de tipo II se basan en una única proteína multifuncional, Cas9, para el paso de interferencia. Cas9 requiere tanto el crRNA como el tracrRNA para funcionar y escindir el ADN utilizando sus dominios de endonucleasa duales tipo HNH y RuvC/RNaseH. En los sistemas de tipo II se requiere el emparejamiento de bases entre el PAM y el genoma del fago. Sin embargo, el PAM se reconoce en la misma cadena que el crRNA (la cadena opuesta a los sistemas de tipo I).
Los sistemas de tipo III, como el tipo I, requieren seis o siete proteínas Cas que se unen a los crRNA. Los sistemas tipo III analizados desde S. solfataricus y P. furiosus ambos se dirigen al ARNm de los fagos en lugar del genoma del ADN del fago, lo que puede hacer que estos sistemas sean excepcionalmente capaces de apuntar a los genomas de fagos basados en ARN. También se descubrió que los sistemas de tipo III se dirigen al ADN además del ARN utilizando una proteína Cas diferente en el complejo, Cas10. Se demostró que la escisión del ADN depende de la transcripción.
El mecanismo para distinguir el ADN propio del extraño durante la interferencia está integrado en los ARNcr y, por lo tanto, probablemente sea común a los tres sistemas. A lo largo del proceso de maduración distintivo de cada tipo principal, todos los ARNcr contienen una secuencia espaciadora y alguna porción de la repetición en uno o ambos extremos. Es la secuencia de repetición parcial la que evita que el sistema CRISPR-Cas se dirija al cromosoma, ya que el emparejamiento de bases más allá de la secuencia espaciadora se señala a sí mismo y previene la escisión del ADN. Las enzimas CRISPR guiadas por ARN se clasifican como enzimas de restricción de tipo V.
Evolución
Se cree que los genes cas en los módulos adaptador y efector del sistema CRISPR-Cas han evolucionado a partir de dos módulos ancestrales diferentes. Un elemento similar a un transposón llamado casposón que codifica la integrasa similar a Cas1 y potencialmente otros componentes del módulo de adaptación se insertó junto al módulo efector ancestral, que probablemente funcionó como un sistema inmunológico innato independiente. Los genes cas1 y cas2 altamente conservados del módulo adaptador evolucionaron a partir del módulo ancestral, mientras que una variedad de genes cas efectores de clase 1 evolucionaron a partir del módulo efector ancestral. La evolución de estos diversos genes cas del módulo efector de clase 1 estuvo guiada por varios mecanismos, como los eventos de duplicación. Por otro lado, cada tipo de módulo efector de clase 2 surgió de inserciones posteriores independientes de elementos genéticos móviles. Estos elementos genéticos móviles reemplazaron los múltiples módulos efectores de genes para crear módulos efectores de un solo gen que producen proteínas grandes que realizan todas las tareas necesarias del módulo efector. Las regiones espaciadoras de los sistemas CRISPR-Cas se toman directamente de elementos genéticos móviles extraños y, por lo tanto, es difícil rastrear su evolución a largo plazo. Se ha descubierto que la evolución no aleatoria de estas regiones espaciadoras depende en gran medida del entorno y de los elementos genéticos móviles extraños particulares que contiene.
CRISPR-Cas puede inmunizar bacterias contra ciertos fagos y así detener la transmisión. Por este motivo, Koonin describió CRISPR-Cas como un mecanismo de herencia lamarckiano. Sin embargo, esto fue cuestionado por un crítico que señaló: "Debemos recordar [a Lamarck] por el bien que contribuyó a la ciencia, no por cosas que se parecen a su teoría sólo superficialmente". De hecho, pensar en CRISPR y otros fenómenos como lamarckianos sólo oscurece la forma simple y elegante en que realmente funciona la evolución. Pero a medida que se han realizado estudios más recientes, se ha hecho evidente que las regiones espaciadoras adquiridas de los sistemas CRISPR-Cas son de hecho una forma de evolución lamarckiana porque son mutaciones genéticas que se adquieren y luego se transmiten. Por otro lado, la evolución de la maquinaria del gen Cas que facilita el sistema evoluciona a través de la evolución darwiniana clásica.
Coevolution
El análisis de secuencias CRISPR reveló la coevolución de los genomas virales y del huésped. Las proteínas Cas9 están altamente enriquecidas en bacterias patógenas y comensales. La regulación genética mediada por CRISPR-Cas puede contribuir a la regulación de genes bacterianos endógenos, particularmente durante la interacción con huéspedes eucariotas. Por ejemplo, Francisella novicida utiliza un ARN pequeño y único asociado a CRISPR-Cas (scaRNA) para reprimir una transcripción endógena que codifica una lipoproteína bacteriana que es fundamental para F. novicida para amortiguar la respuesta del huésped y promover la virulencia.
El modelo básico de la evolución de CRISPR consiste en espaciadores recientemente incorporados que impulsan a los fagos a mutar sus genomas para evitar la respuesta inmune bacteriana, creando diversidad tanto en las poblaciones de fagos como en las del huésped. Para resistir una infección por fagos, la secuencia del espaciador CRISPR debe corresponder perfectamente a la secuencia del gen del fago diana. Los fagos pueden seguir infectando a sus huéspedes. mutaciones puntuales dadas en el espaciador. Se requiere un rigor similar en PAM o la cepa bacteriana seguirá siendo sensible a los fagos.
Tarifas
Un estudio de 124 S. thermophilus mostraron que el 26% de todos los espaciadores eran únicos y que diferentes loci CRISPR mostraban diferentes tasas de adquisición de espaciadores. Algunos loci CRISPR evolucionan más rápidamente que otros, lo que permitió que las cepas evolucionaran más rápidamente. relaciones filogenéticas por determinar. Un análisis genómico comparativo mostró que E. coli y S. enterica evoluciona mucho más lentamente que S. thermophilus. Las cepas de este último que divergieron hace 250.000 años todavía contenían el mismo complemento espacial.
El análisis metagenómico de dos biopelículas de drenaje ácido de minas mostró que una de las CRISPR analizadas contenía eliminaciones extensas y adiciones de espaciadores en comparación con la otra biopelícula, lo que sugiere una mayor actividad/prevalencia de fagos en una comunidad que en la otra. En la cavidad bucal, un estudio temporal determinó que entre el 7% y el 22% de los espaciadores se compartieron durante 17 meses dentro de un individuo, mientras que menos del 2% se compartieron entre individuos.
Del mismo entorno, se rastreó una única cepa utilizando cebadores de PCR específicos de su sistema CRISPR. Los resultados a amplio nivel de presencia/ausencia de espaciadores mostraron una diversidad significativa. Sin embargo, este CRISPR añadió tres espaciadores a lo largo de 17 meses, lo que sugiere que incluso en un entorno con una diversidad CRISPR significativa, algunos loci evolucionan lentamente.
Se analizaron CRISPR a partir de los metagenomas producidos para el Proyecto Microbioma Humano. Aunque la mayoría eran específicas de un sitio corporal, algunas dentro de un sitio corporal son ampliamente compartidas entre los individuos. Uno de estos loci se originó a partir de especies de estreptococos y contenía aproximadamente 15.000 espaciadores, el 50% de los cuales eran únicos. Al igual que los estudios específicos de la cavidad bucal, algunos mostraron poca evolución en el tiempo.
La evolución de CRISPR se estudió en quimiostatos utilizando S. thermophilus para examinar directamente las tasas de adquisición de espaciadores. En una semana, S. Las cepas thermophilus adquirieron hasta tres espaciadores cuando se expusieron a un solo fago. Durante el mismo intervalo, el fago desarrolló polimorfismos de un solo nucleótido que se fijaron en la población, lo que sugiere que la focalización había impedido la replicación del fago en ausencia de estas mutaciones.
Otro S. thermophilus demostró que los fagos pueden infectar y replicarse en huéspedes que tienen un solo espaciador objetivo. Otro más demostró que pueden existir huéspedes sensibles en entornos con títulos elevados de fagos. Los estudios observacionales y de quimiostato sugieren muchos matices para CRISPR y la (co)evolución de los fagos.
Identificación
Los CRISPR están ampliamente distribuidos entre bacterias y arqueas y muestran algunas similitudes de secuencia. Su característica más notable son sus espaciadores repetidos y repeticiones directas. Esta característica hace que los CRISPR sean fácilmente identificables en secuencias largas de ADN, ya que el número de repeticiones disminuye la probabilidad de una coincidencia falsa positiva.
El análisis de CRISPR en datos metagenómicos es más desafiante, ya que los loci CRISPR generalmente no se ensamblan, debido a su naturaleza repetitiva o a través de la variación de tensión, lo que confunde a los algoritmos de ensamblaje. Cuando hay muchos genomas de referencia disponibles, se puede utilizar la reacción en cadena de la polimerasa (PCR) para amplificar las matrices CRISPR y analizar el contenido del espaciador. Sin embargo, este enfoque proporciona información solo para CRISPR específicamente dirigidos y para organismos con suficiente representación en bases de datos públicas para diseñar cebadores de PCR de polimerasa confiables. Se pueden utilizar cebadores degenerados específicos de repetición para amplificar los espaciadores CRISPR directamente a partir de muestras ambientales; Luego, los amplicones que contienen dos o tres espaciadores se pueden ensamblar computacionalmente para reconstruir matrices CRISPR largas.
La alternativa es extraer y reconstruir matrices CRISPR a partir de datos metagenómicos de escopeta. Esto es computacionalmente más difícil, particularmente con las tecnologías de secuenciación de segunda generación (por ejemplo, 454, Illumina), ya que las longitudes de lectura cortas evitan que aparezcan más de dos o tres unidades repetidas en una sola lectura. La identificación CRISPR en lecturas sin procesar se ha logrado mediante identificación puramente de novo o mediante el uso de secuencias repetidas directas en matrices CRISPR parcialmente ensambladas a partir de contigs (segmentos de ADN superpuestos que juntos representan una región de consenso de ADN) y secuencias repetidas directas. de genomas publicados como gancho para identificar repeticiones directas en lecturas individuales.
Uso por fagos
Otra forma que tienen las bacterias de defenderse contra la infección por fagos es tener islas cromosómicas. Un subtipo de islas cromosómicas llamadas islas cromosómicas inducibles por fagos (PICI) se escinde de un cromosoma bacteriano tras la infección por fagos y puede inhibir la replicación de los fagos. Los PICI son inducidos, escindidos, replicados y finalmente empaquetados en pequeñas cápsides por ciertos fagos estafilocócicos templados. Los PICI utilizan varios mecanismos para bloquear la reproducción de fagos. En el primer mecanismo, Ppi codificado por PICI bloquea diferencialmente la maduración de los fagos uniéndose o interactuando específicamente con el fago TerS, bloqueando así la formación del complejo fago TerS/TerL responsable del empaquetado del ADN del fago. En el segundo mecanismo, PICI CpmAB redirige la proteína morfogenética de la cápside del fago para producir el 95% de la cápside del tamaño de SaPI y el ADN del fago puede empaquetar solo 1/3 de su genoma en estas pequeñas cápsides y, por lo tanto, convertirse en un fago no viable. El tercer mecanismo involucra dos proteínas, PtiA y PtiB, que se dirigen a LtrC, que es responsable de la producción de viriones y proteínas de lisis. Este mecanismo de interferencia está modulado por una proteína moduladora, PtiM, que se une a una de las proteínas mediadoras de interferencia, PtiA, y por lo tanto alcanza el nivel requerido de interferencia.
Un estudio demostró que el fago lítico ICP1, que se dirige específicamente al Vibrio cholerae serogrupo O1, ha adquirido un sistema CRISPR-Cas que se dirige a un V. cólera elemento similar al PICI. El sistema tiene 2 loci CRISPR y 9 genes Cas. Parece ser homólogo al sistema I-F que se encuentra en Yersinia pestis. Además, al igual que el sistema bacteriano CRISPR-Cas, ICP1 CRISPR-Cas puede adquirir nuevas secuencias, lo que permite que el fago y el huésped coevolucionen.
Se demostró que ciertos virus de arqueas portan matrices mini-CRISPR que contienen uno o dos espaciadores. Se ha demostrado que los espaciadores dentro de las matrices CRISPR transmitidas por virus se dirigen a otros virus y plásmidos, lo que sugiere que las matrices mini-CRISPR representan un mecanismo de exclusión de superinfección heterotípica y participan en conflictos intervirales.
Aplicaciones
Edición de genes CRISPR
La tecnología CRISPR se ha aplicado en las industrias alimentaria y agrícola para diseñar cultivos probióticos e inmunizar cultivos industriales (para yogur, por ejemplo) contra infecciones. También se utiliza en cultivos para mejorar el rendimiento, la tolerancia a la sequía y el valor nutricional. La edición de genes CRISPR también se ha convertido en una herramienta fantástica para la investigación genética científica. La amplificación y el "knock-out" de productos genéticos es una forma confiable de identificar genes de interés para el desarrollo farmacéutico o simplemente de comprender mejor las complejidades que se encuentran en cualquier genoma.
A finales de 2014, se habían publicado unos 1.000 artículos de investigación que mencionaban CRISPR. La tecnología se había utilizado para inactivar funcionalmente genes en líneas celulares y células humanas, para estudiar Candida albicans, para modificar levaduras utilizadas para producir biocombustibles y modificar genéticamente cepas de cultivos. Hsu y sus colegas afirman que la capacidad de manipular las secuencias genéticas permite realizar ingeniería inversa que puede afectar positivamente la producción de biocombustibles. CRISPR también se puede utilizar para editar genéticamente mosquitos para que no puedan transmitir enfermedades como la malaria. Recientemente se han utilizado enfoques basados en CRISPR que utilizan Cas12a en la modificación exitosa de una amplia cantidad de especies de plantas.
En julio de 2019, se utilizó CRISPR para tratar experimentalmente a un paciente con un trastorno genético. La paciente era una mujer de 34 años con anemia falciforme.
En febrero de 2020, se lograron avances en los tratamientos contra el VIH: entre el 60% y el 80% del ADN viral integrado se eliminó en ratones y algunos quedaron completamente libres del virus después de ediciones que involucraron tanto LASER ART, una nueva terapia antirretroviral, como CRISPR. .
En marzo de 2020, se inyectó virus modificado con CRISPR en el ojo de un paciente en un intento de tratar la amaurosis congénita de Leber.
En el futuro, la edición de genes CRISPR podría utilizarse para crear nuevas especies o revivir especies extintas a partir de especies estrechamente relacionadas.
Las reevaluaciones basadas en CRISPR de afirmaciones sobre relaciones entre genes y enfermedades han llevado al descubrimiento de anomalías potencialmente importantes.
En julio de 2021, se utilizó la edición genética CRISPR de hiPSC para estudiar el papel de las proteínas MBNL asociadas con la DM1.
La técnica CRISPR tiene una respuesta positiva al trabajar con diferentes tipos de células y tejidos y, por lo tanto, con diferentes trastornos como el sistema nervioso, el sistema circulatorio, las células madre, los trastornos sanguíneos y la degeneración muscular. Esta herramienta ha generado enfoques avanzados tanto en sistemas terapéuticos como biomédicos y algunas de sus aplicaciones se analizan a continuación.
1.1 β-Hemoglobinopatías
Esta enfermedad pertenece a los trastornos genéticos causados por una mutación que se produce en la estructura de la hemoglobina o por la sustitución de diferentes aminoácidos en las cadenas de globina. Debido a esto, los glóbulos rojos (RBC) causan una serie de obstáculos como insuficiencia cardíaca, obstrucción de los vasos sanguíneos, defectos en el crecimiento y problemas ópticos. Para rehabilitar las β-hemoglobinopatías, las células multipotentes del paciente se transfieren a un modelo de ratón para estudiar la tasa de terapia génica ex vivo, lo que da como resultado la expresión del ARNm y el gen que se rectifica. Curiosamente, la vida media de los glóbulos rojos también aumentó.
1.2 Hemofilia
Es una pérdida de función en la sangre donde los factores de coagulación no funcionan correctamente. Hay dos tipos, hemofilia A y hemofilia B. Mediante el uso de CRISPR-Cas9, se inserta un vector en las bacterias. El vector utilizado es un vector adenoviral que ayuda en la corrección de genes. Sin duda, CRISPR ha dado esperanzas para el tratamiento de la hemofilia al corregir los genes.
1.3 Trastornos neurológicos
CRISPR se utiliza para suprimir las mutaciones que causan la ganancia de función y también repara las mutaciones con pérdida de funciones mediante la edición de genes en trastornos neurológicos. La herramienta de edición de genes estableció un punto de apoyo en la aplicación in vivo para la asimilación de vías moleculares.
1.4 Ceguera
Los trastornos oculares se convirtieron en un impedimento más para que los médicos trataran a las víctimas. Además, el tejido de la retina presente en el ojo está libre de respuesta inmune del cuerpo. Las enfermedades oculares más frecuentes en todo el mundo son las cataratas y la retinitis pigmentosa (RP). Estos son causados por una mutación sin sentido en la cadena alfa que conduce a la ceguera permanente. El enfoque de CRISPR es empaquetar el gen que codifica la proteína retiniana y editar el genoma, lo que da como resultado una buena visión.
1.5 Enfermedades cardiovasculares
La tecnología CRISPR actúa de manera más eficiente contra las enfermedades relacionadas con el corazón. Debido a la deposición de colesterol en las paredes de las arterias, se produce un bloqueo del flujo sanguíneo. Esto es causado por una mutación en los receptores de colesterol de lipoproteínas de baja densidad (LDLC), que da como resultado la liberación de colesterol a la sangre en niveles más altos. Esto puede tratarse mediante la eliminación del par de bases en el exón 4 del receptor LDLC. Esta es una mutación sin sentido.
Aplicaciones de CRISPR en agricultura
La aplicación de CRISPR en plantas se logró con éxito en 2013. CRISPR Cas9 se ha convertido en un instrumento influyente en la edición de genomas en cultivos. Dejó su huella en los sistemas de cría actuales,
2.1 Impulsar el rendimiento
Para lograr un alto rendimiento en los cereales, se modifica el equilibrio de la citoquinina. La citoquinina oxidasa/deshidrogenasa (CKX) es una enzima, por lo que el gen que codifica esta enzima fue eliminado para producir más rendimiento.
2.2 Mejora de la calidad
Los cereales tienen una gran cantidad de polisacárido de amilosa. Para disminuir el contenido de amilosa, se usa CRISPR para alterar los aminoácidos, lo que conduce a una baja producción de sacárido. Además, el trigo contiene proteínas de gluten, por lo que algunos de ellos son intolerantes al gluten y provocan un trastorno llamado enfermedad celíaca. La herramienta de edición de genes se dirige a los genes del gluten, lo que da como resultado una baja producción de gluten en el trigo.
2.3 Resistencia a las enfermedades
El estrés biótico de las plantas se puede reducir mediante el uso de herramientas CRISPR. Las infecciones bacterianas causadas en el arroz provocan la activación de la transcripción de genes, cuyos productos son susceptibles a enfermedades y mediante el uso de CRISPR los científicos pudieron generar líneas de resistencia.
Aplicaciones generales de CRISPR
3.1 Terapia génica
El total de trastornos genéticos descubiertos hasta ahora son alrededor de 6000. La mayoría de ellos no cuentan con tratamiento hasta la fecha. La función de la terapia génica es sustituir los genes defectuosos por ADN exógeno o editar la secuencia mutada para reflejar la secuencia genética sana. Esta terapia tuvo un gran impacto en la biotecnología médica.
3.2 Edición básica
Son dos tipos de ediciones base:
El editor base de citidina es una terapia novedosa en la que la citidina (C) cambia a timidina (T).
Editor de bases de adenina (ABE), en este hay un cambio de complementos de bases de adenina (A) a Guanina (G).
Las mutaciones se instalaron directamente en el ADN celular, por lo que no se requiere la plantilla del donante. Las ediciones base solo pueden editar mutaciones puntuales y además solo pueden corregir mutaciones de hasta cuatro puntos. Entonces, para resolver este problema, el sistema CRISPR ha introducido una nueva técnica conocida como fusión Cas9 para ampliar el nivel de edición del genoma.
3.3 Silenciamiento y activación de genes
Además, la proteína CRISPR Cas9 puede modular genes activándolos o silenciándolos en función de genes de interés. Existe una nucleasa llamada dCas9 (endonucleasa) que se utiliza para silenciar o activar la expresión de genes.
Limitaciones en las aplicaciones de CRISPR
Los investigadores se enfrentan a muchos desafíos en la edición de genes. Los principales obstáculos que surgen en las aplicaciones clínicas son las cuestiones éticas y el sistema de transporte al sitio de destino. Como las unidades del sistema CRISPR se toman de bacterias, cuando se transfieren a las células huésped se produce una respuesta inmune contra ellas. Se utilizan vectores virales físicos y químicos como vehículos para llevar el complejo al huésped. Debido a esto, surgen muchas complicaciones, como daño celular que conduce a la muerte celular. En el caso de los vectores virales, la capacidad del virus es pequeña y la proteína Cas9 es grande. Entonces, para superar estos nuevos métodos, se desarrollaron nuevos métodos en los que se toman cepas más pequeñas de Cas9 de bacterias. Aún queda mucho trabajo por hacer para mejorar el sistema.
CRISPR como herramienta de diagnóstico

Las nucleasas asociadas a CRISPR han demostrado ser útiles como herramienta para pruebas moleculares debido a su capacidad para apuntar específicamente a secuencias de ácido nucleico en un fondo elevado de secuencias no objetivo. En 2016, la nucleasa Cas9 se utilizó para agotar secuencias de nucleótidos no deseadas en bibliotecas de secuenciación de próxima generación y solo requirió 250 picogramos de entrada inicial de ARN. A partir de 2017, las nucleasas asociadas a CRISPR también se utilizaron para pruebas de diagnóstico directo de ácidos nucleicos, hasta la sensibilidad de una sola molécula. La diversidad CRISPR se utiliza como objetivo de análisis para discernir la filogenia y la diversidad en bacterias, como en las xantomonas, por Martins et al., 2019. Detecciones tempranas de patógenos vegetales mediante tipificación molecular de los patógenos Los CRISPR se pueden utilizar en la agricultura, como lo demuestran Shen et al., 2020.
Al acoplar el diagnóstico basado en CRISPR a procesos enzimáticos adicionales, es posible la detección de moléculas más allá de los ácidos nucleicos. Un ejemplo de una tecnología acoplada es el perfilado de transcripción in vitro (SPRINT) basado en SHERLOCK. SPRINT se puede utilizar para detectar una variedad de sustancias, como metabolitos en muestras de pacientes o contaminantes en muestras ambientales, con alto rendimiento o con dispositivos portátiles en el punto de atención. También se están explorando plataformas CRISPR-Cas para la detección e inactivación del SARS-CoV-2, el virus que causa el COVID-19. Se han identificado dos pruebas de diagnóstico integral diferentes, AIOD-CRISPR y la prueba SHERLOCK para el SARS-CoV-2. La prueba SHERLOCK se basa en un ARN de reportero de prensa marcado con fluorescencia que tiene la capacidad de identificar 10 copias por microlitro. AIOD-CRISPR ayuda con una detección visual sólida y altamente sensible del ácido nucleico viral.