
Corteza cerebral
La corteza cerebral, también conocida como manto cerebral, es la capa externa de tejido neural del cerebro del cerebro en humanos y otros mamíferos. La corteza cerebral consiste principalmente en la neocorteza de seis capas, y solo el 10% consiste en alocorteza. Está separado en dos cortezas por la fisura longitudinal que divide el cerebro en los hemisferios cerebrales izquierdo y derecho. Los dos hemisferios están unidos debajo de la corteza por el cuerpo calloso. La corteza cerebral es el sitio más grande de integración neuronal en el sistema nervioso central. Desempeña un papel clave en la atención, la percepción, la conciencia, el pensamiento, la memoria, el lenguaje y la conciencia. La corteza cerebral es parte del cerebro responsable de la cognición.
En la mayoría de los mamíferos, aparte de los pequeños mamíferos que tienen cerebros pequeños, la corteza cerebral está plegada, proporcionando una mayor superficie en el volumen confinado del cráneo. Además de minimizar el volumen cerebral y craneal, el plegamiento cortical es crucial para el circuito cerebral y su organización funcional. En los mamíferos con cerebro pequeño no hay plegamiento y la corteza es lisa.
Un pliegue o cresta en la corteza se denomina circunvolución (plural gyri) y un surco se denomina surco (plural sulci). Estas circunvoluciones superficiales aparecen durante el desarrollo fetal y continúan madurando después del nacimiento a través del proceso de girificación. En el cerebro humano, la mayor parte de la corteza cerebral no es visible desde el exterior, sino enterrada en los surcos. Los surcos y circunvoluciones principales marcan las divisiones del cerebro en los lóbulos del cerebro. Los cuatro lóbulos principales son los lóbulos frontal, parietal, occipital y temporal. Otros lóbulos son el lóbulo límbico y la corteza insular a menudo denominada lóbulo insular.
Hay entre 14 y 16 mil millones de neuronas en la corteza cerebral humana. Estos se organizan en capas corticales horizontales y radialmente en columnas y minicolumnas corticales. Las áreas corticales tienen funciones específicas como el movimiento en la corteza motora y la vista en la corteza visual. La corteza visual se encuentra en el lóbulo occipital.
Estructura

La corteza cerebral es la cubierta exterior de las superficies de los hemisferios cerebrales y está plegada en picos llamados circunvoluciones y surcos llamados surcos. En el cerebro humano tiene entre dos y tres o cuatro milímetros de espesor, y constituye el 40 por ciento de la masa cerebral. El 90 por ciento de la corteza cerebral es la neocorteza de seis capas y el otro 10 por ciento está formado por alocorteza. Hay entre 14 y 16 mil millones de neuronas en la corteza, y están organizadas radialmente en columnas corticales y minicolumnas, en las capas organizadas horizontalmente de la corteza.
La neocorteza se puede separar en diferentes regiones de la corteza conocidas en plural como cortezas, e incluyen la corteza motora y la corteza visual. Aproximadamente dos tercios de la superficie cortical están enterrados en los surcos y la corteza insular está completamente oculta. La corteza es más gruesa sobre la parte superior de una circunvolución y más delgada en la parte inferior de un surco.
Pliegues
La corteza cerebral está plegada de tal forma que permite que una gran superficie de tejido neural quepa dentro de los confines del neurocráneo. Cuando se despliega en el ser humano, cada corteza hemisférica tiene una superficie total de aproximadamente 0,12 metros cuadrados (1,3 pies cuadrados). El pliegue se aleja de la superficie del cerebro y también está presente en la superficie medial de cada hemisferio dentro de la fisura longitudinal. La mayoría de los mamíferos tienen una corteza cerebral que se enreda con los picos conocidos como circunvoluciones y los valles o surcos conocidos como surcos. Algunos pequeños mamíferos, incluidos algunos pequeños roedores, tienen superficies cerebrales lisas sin girificación.
Lóbulos
Los surcos y circunvoluciones más grandes marcan las divisiones de la corteza del cerebro en los lóbulos del cerebro. Hay cuatro lóbulos principales: el lóbulo frontal, el lóbulo parietal, el lóbulo temporal y el lóbulo occipital. La corteza insular a menudo se incluye como el lóbulo insular. El lóbulo límbico es un borde de la corteza en el lado medial de cada hemisferio y también se incluye a menudo. También se describen tres lóbulos del cerebro: el lóbulo paracentral, el lóbulo parietal superior y el lóbulo parietal inferior.
Espesor
Para las especies de mamíferos, los cerebros más grandes (en términos absolutos, no solo en relación con el tamaño del cuerpo) tienden a tener cortezas más gruesas. Los mamíferos más pequeños, como las musarañas, tienen un grosor neocortical de unos 0,5 mm; los que tienen los cerebros más grandes, como los humanos y los rorcuales comunes, tienen un grosor de 2 a 4 mm. Existe una relación aproximadamente logarítmica entre el peso del cerebro y el grosor cortical. La resonancia magnética del cerebro (IRM) permite obtener una medida del grosor de la corteza cerebral humana y relacionarla con otras medidas. El grosor de las diferentes áreas corticales varía pero, en general, la corteza sensorial es más delgada que la corteza motora. Un estudio ha encontrado una asociación positiva entre el grosor cortical y la inteligencia. Otro estudio ha encontrado que la corteza somatosensorial es más gruesa en pacientes con migraña, aunque no se sabe si esto es el resultado de los ataques de migraña, la causa de los mismos o si ambos son el resultado de una causa compartida. Un estudio posterior que utilizó una población de pacientes más grande no informa cambios en el grosor cortical en pacientes con migraña. Un trastorno genético de la corteza cerebral, por el cual la disminución del plegamiento en ciertas áreas da como resultado un microgiro, donde hay cuatro capas en lugar de seis, en algunos casos se considera que está relacionado con la dislexia.
Capas de neocorteza



La neocorteza está formada por seis capas, numeradas de la I a la VI, desde la capa más externa I, cerca de la piamadre, hasta la capa más interna VI, cerca de la sustancia blanca subyacente. Cada capa cortical tiene una distribución característica de diferentes neuronas y sus conexiones con otras regiones corticales y subcorticales. Hay conexiones directas entre diferentes áreas corticales y conexiones indirectas a través del tálamo.
Uno de los ejemplos más claros de capas corticales es la línea de Gennari en la corteza visual primaria. Se trata de una banda de tejido más blanco que se observa a simple vista en el surco calcarino del lóbulo occipital. La línea de Gennari está compuesta por axones que traen información visual desde el tálamo a la capa IV de la corteza visual.
La tinción de secciones transversales de la corteza para revelar la posición de los cuerpos celulares neuronales y los tractos axónicos intracorticales permitió a los neuroanatomistas de principios del siglo XX producir una descripción detallada de la estructura laminar de la corteza en especies diferentes. El trabajo de Korbinian Brodmann (1909) estableció que la neocorteza de los mamíferos se divide consistentemente en seis capas.
Capa I
La capa I es la capa molecular y contiene pocas neuronas dispersas, incluidas las neuronas de rosa mosqueta GABAérgicas. La capa I consta en gran parte de extensiones de penachos dendríticos apicales de neuronas piramidales y axones orientados horizontalmente, así como células gliales. Durante el desarrollo, las células de Cajal-Retzius y las células de la capa granular subpial están presentes en esta capa. Además, aquí se pueden encontrar algunas células estrelladas espinosas. Se cree que las entradas a los mechones apicales son cruciales para las interacciones de retroalimentación en la corteza cerebral involucradas en el aprendizaje asociativo y la atención. Si bien alguna vez se pensó que la entrada a la capa I provenía de la corteza misma, ahora se sabe que la capa I a través del manto de la corteza cerebral recibe una entrada sustancial de matriz o células del tálamo tipo M (en contraste a núcleo o tipo C que van a la capa IV).
Capa II
La capa II, la capa granular externa, contiene pequeñas neuronas piramidales y numerosas neuronas estrelladas.
Capa III
La capa III, la capa piramidal externa, contiene predominantemente neuronas piramidales de tamaño pequeño y mediano, así como neuronas no piramidales con axones intracorticales orientados verticalmente; las capas I a III son el objetivo principal de las fibras aferentes corticocorticales interhemisféricas, y la capa III es la fuente principal de las fibras eferentes corticocorticales.
Capa IV
La capa IV, la capa granular interna, contiene diferentes tipos de células estrelladas y piramidales, y es el objetivo principal de los aferentes talamocorticales de las neuronas tipo C del tálamo (tipo central), así como de los aferentes corticocorticales intrahemisféricos. Las capas por encima de la capa IV también se denominan capas supragranulares (capas I-III), mientras que las capas inferiores se denominan capas infragranulares (capas V y VI).
Capa V
La capa V, la capa piramidal interna, contiene neuronas piramidales grandes. Los axones de estos salen de la corteza y se conectan con estructuras subcorticales, incluidos los ganglios basales. En la corteza motora primaria del lóbulo frontal, la capa V contiene células piramidales gigantes llamadas células de Betz, cuyos axones viajan a través de la cápsula interna, el tronco encefálico y la médula espinal formando el tracto corticoespinal, que es la vía principal para el control motor voluntario..
Capa VI
La capa VI, la capa polimórfica o multiforme, contiene pocas neuronas piramidales grandes y muchas neuronas piramidales y multiformes pequeñas en forma de huso; la capa VI envía fibras eferentes al tálamo, estableciendo una interconexión recíproca muy precisa entre la corteza y el tálamo. Es decir, las neuronas de la capa VI de una columna cortical se conectan con las neuronas del tálamo que proporcionan información a la misma columna cortical. Estas conexiones son excitatorias e inhibitorias. Las neuronas envían fibras excitatorias a las neuronas del tálamo y también envían colaterales al núcleo reticular talámico que inhiben estas mismas neuronas del tálamo o las adyacentes. Una teoría es que debido a que la salida inhibidora se reduce por la entrada colinérgica a la corteza cerebral, esto proporciona al tronco encefálico un "control de ganancia ajustable para el relé de entradas lemniscales".
Columnas
Las capas corticales no están simplemente apiladas unas sobre otras; existen conexiones características entre diferentes capas y tipos neuronales, que abarcan todo el espesor de la corteza. Estos microcircuitos corticales se agrupan en columnas y minicolumnas corticales. Se ha propuesto que las minicolumnas son las unidades funcionales básicas de la corteza. En 1957, Vernon Mountcastle demostró que las propiedades funcionales de la corteza cambian abruptamente entre puntos lateralmente adyacentes; sin embargo, son continuas en la dirección perpendicular a la superficie. Trabajos posteriores han proporcionado evidencia de la presencia de columnas corticales funcionalmente distintas en la corteza visual (Hubel y Wiesel, 1959), la corteza auditiva, y corteza asociativa.
Las áreas corticales que carecen de una capa IV se denominan agranulares. Las áreas corticales que tienen solo una capa IV rudimentaria se denominan disgranulares. El procesamiento de la información dentro de cada capa está determinado por diferentes dinámicas temporales, ya que en las capas II/III tiene una oscilación lenta de 2 Hz, mientras que en la capa V tiene una oscilación rápida de 10 a 15 Hz.
Tipos de corteza
Según las diferencias en la organización laminar, la corteza cerebral se puede clasificar en dos tipos, el área grande de neocorteza que tiene seis capas de células y el área mucho más pequeña de alocorteza que tiene tres o cuatro capas:
- El neocortex también se conoce como el isocortex o el neopalio y es la parte de la corteza cerebral madura con seis capas distintas. Ejemplos de áreas neocorticales incluyen la corteza motora primaria granular, y la corteza visual primaria estriada. El neocortex tiene dos subtipos, el verdadero isocortex y el proisocortex que es una región de transición entre el isocortex y las regiones del periallocortex.
- El alocortex es la parte de la corteza cerebral con tres o cuatro capas, y tiene tres subtipos, el paleocortex con tres laminas corticales, el archicortex que tiene cuatro o cinco, y un área de transición adyacente al alocortex, el periallocortex. Ejemplos de alocortex son la corteza olfativa y el hipocampo.
Hay un área de transición entre la neocorteza y la alocorteza llamada corteza paralímbica, donde se fusionan las capas 2, 3 y 4. Esta área incorpora la proisocorteza de la neocorteza y la periallocorteza de la alocorteza. Además, la corteza cerebral se puede clasificar en cuatro lóbulos: el lóbulo frontal, el lóbulo temporal, el lóbulo parietal y el lóbulo occipital, denominados así por los huesos del cráneo que los recubren.
Abastecimiento y drenaje de sangre

El suministro de sangre a la corteza cerebral es parte de la circulación cerebral. Las arterias cerebrales suministran la sangre que irriga el cerebro. Esta sangre arterial transporta oxígeno, glucosa y otros nutrientes a la corteza. Las venas cerebrales drenan la sangre desoxigenada y los desechos metabólicos, incluido el dióxido de carbono, de regreso al corazón.
Las principales arterias que irrigan la corteza son la arteria cerebral anterior, la arteria cerebral media y la arteria cerebral posterior. La arteria cerebral anterior irriga las porciones anteriores del cerebro, incluida la mayor parte del lóbulo frontal. La arteria cerebral media irriga los lóbulos parietales, los lóbulos temporales y partes de los lóbulos occipitales. La arteria cerebral media se divide en dos ramas para irrigar los hemisferios izquierdo y derecho, donde se ramifican aún más. La arteria cerebral posterior irriga los lóbulos occipitales.
El círculo de Willis es el principal sistema sanguíneo que se ocupa del suministro de sangre en el cerebro y la corteza cerebral.

Desarrollo
El desarrollo prenatal de la corteza cerebral es un proceso complejo y finamente sintonizado llamado corticogénesis, influenciado por la interacción entre los genes y el medio ambiente.
Tubo neural
La corteza cerebral se desarrolla a partir de la parte más anterior, la región del cerebro anterior, del tubo neural. La placa neural se pliega y se cierra para formar el tubo neural. A partir de la cavidad interior del tubo neural se desarrolla el sistema ventricular y, a partir de las células neuroepiteliales de sus paredes, las neuronas y glía del sistema nervioso. La parte más anterior (frontal o craneal) de la placa neural, el prosencéfalo, que es evidente antes de que comience la neurulación, da origen a los hemisferios cerebrales y posteriormente a la corteza.
Desarrollo de neuronas corticales
Las neuronas corticales se generan dentro de la zona ventricular, junto a los ventrículos. Al principio, esta zona contiene células madre neurales, que hacen la transición a células gliales radiales: células progenitoras, que se dividen para producir células gliales y neuronas.
Glía radial

La corteza cerebral está compuesta por una población heterogénea de células que dan lugar a diferentes tipos de células. La mayoría de estas células se derivan de la migración de la glía radial que forma los diferentes tipos celulares del neocórtex y es un período asociado a un aumento de la neurogénesis. De manera similar, el proceso de neurogénesis regula la laminación para formar las diferentes capas de la corteza. Durante este proceso hay un aumento en la restricción del destino celular que comienza con progenitores más tempranos que dan lugar a cualquier tipo de célula en la corteza y progenitores posteriores que dan lugar solo a neuronas de capas superficiales. Este destino celular diferencial crea una topografía de adentro hacia afuera en la corteza con neuronas más jóvenes en capas superficiales y neuronas más viejas en capas más profundas. Además, las neuronas laminares se detienen en la fase S o G2 para dar una distinción fina entre las diferentes capas corticales. La diferenciación laminar no está completamente completa hasta después del nacimiento, ya que durante el desarrollo, las neuronas laminares aún son sensibles a señales extrínsecas y señales ambientales.
Aunque la mayoría de las células que componen la corteza se derivan localmente de la glía radial, existe un subconjunto de la población de neuronas que migran desde otras regiones. La glía radial da lugar a neuronas que tienen forma piramidal y usan glutamato como neurotransmisor, sin embargo, estas células migratorias aportan neuronas que tienen forma estrellada y usan GABA como su principal neurotransmisor. Estas neuronas GABAérgicas son generadas por células progenitoras en la eminencia ganglionar medial (MGE) que migran tangencialmente a la corteza a través de la zona subventricular. Esta migración de neuronas GABAérgicas es particularmente importante ya que los receptores GABA son excitadores durante el desarrollo. Esta excitación es impulsada principalmente por el flujo de iones de cloruro a través del receptor GABA; sin embargo, en los adultos, las concentraciones de cloruro cambian y provocan un flujo de cloruro hacia el interior que hiperpolariza las neuronas postsinápticas. Las fibras gliales producidas en las primeras divisiones de las células progenitoras están orientadas radialmente, abarcando el grosor de la corteza desde la zona ventricular hasta la superficie externa pial, y proporcionan andamiaje para la migración de neuronas hacia el exterior desde la zona ventricular.
Al nacer, hay muy pocas dendritas presentes en el cuerpo celular de la neurona cortical y el axón no está desarrollado. Durante el primer año de vida, el número de dendritas aumenta drásticamente, de modo que pueden albergar hasta cien mil conexiones sinápticas con otras neuronas. El axón puede desarrollarse para extenderse mucho desde el cuerpo celular.
División asimétrica
Las primeras divisiones de las células progenitoras son simétricas, lo que duplica el número total de células progenitoras en cada ciclo mitótico. Luego, algunas células progenitoras comienzan a dividirse asimétricamente, produciendo una célula posmitótica que migra a lo largo de las fibras gliales radiales, saliendo de la zona ventricular, y una célula progenitora, que continúa dividiéndose hasta el final del desarrollo, cuando se diferencia en una célula glial o una célula ependimaria. A medida que se alarga la fase G1 de la mitosis, en lo que se considera un alargamiento selectivo del ciclo celular, las neuronas recién nacidas migran a capas más superficiales de la corteza. Las células hijas migratorias se convierten en las células piramidales de la corteza cerebral. El proceso de desarrollo está ordenado en el tiempo y regulado por cientos de genes y mecanismos reguladores epigenéticos.
Organización de capas

La estructura en capas de la corteza cerebral madura se forma durante el desarrollo. Las primeras neuronas piramidales generadas migran fuera de la zona ventricular y de la zona subventricular, junto con las neuronas de Cajal-Retzius productoras de reelina, de la preplaca. A continuación, una cohorte de neuronas que migran hacia el medio de la preplaca divide esta capa transitoria en la zona marginal superficial, que se convertirá en la capa I de la neocorteza madura, y la subplaca, formando una capa intermedia llamada placa cortical. Estas células formarán las capas profundas de la corteza madura, las capas cinco y seis. Las neuronas nacidas más tarde migran radialmente hacia la placa cortical más allá de las neuronas de la capa profunda y se convierten en las capas superiores (dos a cuatro). Por lo tanto, las capas de la corteza se crean en un orden de adentro hacia afuera. La única excepción a esta secuencia de neurogénesis de adentro hacia afuera ocurre en la capa I de los primates, en la que, a diferencia de los roedores, la neurogénesis continúa durante todo el período de corticogénesis.
Patrones corticales

El mapa de áreas corticales funcionales, que incluyen la corteza visual y motora primaria, se origina a partir de un 'protomapa', que está regulado por señales moleculares como el factor de crecimiento de fibroblastos FGF8 en las primeras etapas del desarrollo embrionario. Estas señales regulan el tamaño, la forma y la posición de las áreas corticales en la superficie del primordio cortical, en parte mediante la regulación de los gradientes de expresión del factor de transcripción, a través de un proceso llamado patrón cortical. Los ejemplos de tales factores de transcripción incluyen los genes EMX2 y PAX6. Juntos, ambos factores de transcripción forman un gradiente de expresión opuesto. Pax6 se expresa mucho en el polo lateral rostral, mientras que Emx2 se expresa mucho en el polo caudomedial. El establecimiento de este gradiente es importante para un correcto desarrollo. Por ejemplo, las mutaciones en Pax6 pueden hacer que los niveles de expresión de Emx2 se expandan fuera de su dominio de expresión normal, lo que en última instancia conduciría a una expansión de las áreas normalmente derivadas de la corteza caudal medial, como la corteza visual. Por el contrario, si se producen mutaciones en Emx2, puede hacer que el dominio que expresa Pax6 se expanda y provoque el agrandamiento de las regiones corticales motoras y frontales. Por lo tanto, los investigadores creen que gradientes y centros de señalización similares próximos a la corteza podrían contribuir a la expresión regional de estos factores de transcripción. Dos señales de patrones muy bien estudiadas para la corteza incluyen FGF y ácido retinoico. Si los FGF se expresan incorrectamente en diferentes áreas de la corteza en desarrollo, se interrumpe el patrón cortical. Específicamente, cuando Fgf8 aumenta en el polo anterior, Emx2 disminuye y se produce un desplazamiento caudal en la región cortical. En última instancia, esto provoca una expansión de las regiones rostrales. Por lo tanto, Fgf8 y otros FGF desempeñan un papel en la regulación de la expresión de Emx2 y Pax6 y representan cómo la corteza cerebral puede especializarse para diferentes funciones.
La rápida expansión del área de la superficie cortical está regulada por la cantidad de autorrenovación de las células gliales radiales y está regulada en parte por los genes FGF y Notch. Durante el período de neurogénesis cortical y formación de capas, muchos mamíferos superiores inician el proceso de girificación, que genera los pliegues característicos de la corteza cerebral. La girificación está regulada por una proteína Trnp1 asociada al ADN y por la señalización de FGF y SHH
Evolución
De todas las diferentes regiones del cerebro, la corteza cerebral muestra la variación evolutiva más grande y ha evolucionado más recientemente. En contraste con el circuito altamente conservado del bulbo raquídeo, por ejemplo, que cumple funciones críticas como la regulación del ritmo cardíaco y respiratorio, muchas áreas de la corteza cerebral no son estrictamente necesarias para la supervivencia. Por lo tanto, la evolución de la corteza cerebral ha visto el advenimiento y la modificación de nuevas áreas funcionales, en particular áreas de asociación que no reciben información directamente del exterior de la corteza.
Una teoría clave de la evolución cortical está incorporada en la hipótesis de la unidad radial y la hipótesis del protomapa relacionada, propuestas por primera vez por Rakic. Esta teoría establece que las nuevas áreas corticales se forman mediante la adición de nuevas unidades radiales, lo que se logra a nivel de células madre. La hipótesis del protomapa establece que la identidad celular y molecular y las características de las neuronas en cada área cortical están especificadas por células madre corticales, conocidas como células gliales radiales, en un mapa primordial. Este mapa está controlado por proteínas de señalización secretadas y factores de transcripción aguas abajo.
Función

Conexiones
La corteza cerebral está conectada a varias estructuras subcorticales como el tálamo y los ganglios basales, les envía información a través de conexiones eferentes y recibe información de ellos a través de conexiones aferentes. La mayor parte de la información sensorial se envía a la corteza cerebral a través del tálamo. La información olfativa, sin embargo, pasa a través del bulbo olfativo a la corteza olfativa (corteza piriforme). La mayoría de las conexiones son de un área de la corteza a otra, en lugar de áreas subcorticales; Braitenberg y Schüz (1998) afirman que en las áreas sensoriales primarias, en el nivel cortical donde terminan las fibras de entrada, hasta el 20 % de las sinapsis son suministradas por aferentes extracorticales, pero que en otras áreas y otras capas es probable que el porcentaje sea mucho mayor. más bajo.
Áreas corticales
Toda la corteza cerebral se dividió en 52 áreas diferentes en una presentación inicial de Korbinian Brodmann. Estas áreas, conocidas como áreas de Brodmann, se basan en su citoarquitectura pero también se relacionan con varias funciones. Un ejemplo es el área 17 de Brodmann, que es la corteza visual primaria.
En términos más generales, la corteza se suele describir como compuesta de tres partes: áreas sensoriales, motoras y de asociación.
Áreas sensoriales


Las áreas sensoriales son las áreas corticales que reciben y procesan la información de los sentidos. Las partes de la corteza que reciben información sensorial del tálamo se denominan áreas sensoriales primarias. Los sentidos de la vista, el oído y el tacto son atendidos por la corteza visual primaria, la corteza auditiva primaria y la corteza somatosensorial primaria, respectivamente. En general, los dos hemisferios reciben información del lado opuesto (contralateral) del cuerpo. Por ejemplo, la corteza somatosensorial primaria derecha recibe información de las extremidades izquierdas y la corteza visual derecha recibe información del campo visual izquierdo. La organización de los mapas sensoriales en la corteza refleja la del órgano sensorial correspondiente, en lo que se conoce como mapa topográfico. Los puntos vecinos en la corteza visual primaria, por ejemplo, corresponden a puntos vecinos en la retina. Este mapa topográfico se llama mapa retinotópico. Del mismo modo, existe un mapa tonotópico en la corteza auditiva primaria y un mapa somatotópico en la corteza sensorial primaria. Este último mapa topográfico del cuerpo en la circunvolución central posterior se ha ilustrado como una representación humana deformada, el homúnculo somatosensorial, donde el tamaño de las diferentes partes del cuerpo refleja la densidad relativa de su inervación. Las áreas con mucha inervación sensorial, como las yemas de los dedos y los labios, requieren más área cortical para procesar sensaciones más finas.
Áreas motoras
Las áreas motoras están ubicadas en ambos hemisferios de la corteza. Las áreas motoras están muy relacionadas con el control de los movimientos voluntarios, especialmente los movimientos fragmentados finos que realiza la mano. La mitad derecha del área motora controla el lado izquierdo del cuerpo y viceversa.
Dos áreas de la corteza se conocen comúnmente como motoras:
- Corteza motora primaria, que ejecutadas Movimientos voluntarios
- Áreas de motor suplementarias y corteza premotor, que seleccionar Movimientos voluntarios.
Además, se han descrito funciones motoras para:
- Posterior corteza parietal, que guía movimientos voluntarios en el espacio
- Corteza prefrontal Dorsolateral, que decide qué movimientos voluntarios deben hacer según instrucciones, reglas y pensamientos autogenerados.
Justo debajo de la corteza cerebral hay masas subcorticales interconectadas de materia gris llamadas ganglios basales (o núcleos). Los ganglios basales reciben información de la sustancia negra del mesencéfalo y de las áreas motoras de la corteza cerebral y envían señales a ambos lugares. Intervienen en el control motor. Se encuentran laterales al tálamo. Los componentes principales de los ganglios basales son el núcleo caudado, el putamen, el globo pálido, la sustancia negra, el núcleo accumbens y el núcleo subtalámico. El putamen y el globo pálido también se conocen colectivamente como núcleo lentiforme, porque juntos forman un cuerpo en forma de lente. El putamen y el núcleo caudado también se denominan colectivamente cuerpo estriado debido a su apariencia rayada.
Áreas de asociación

Las áreas de asociación son las partes de la corteza cerebral que no pertenecen a las regiones primarias. Funcionan para producir una experiencia perceptiva significativa del mundo, nos permiten interactuar de manera efectiva y respaldan el pensamiento y el lenguaje abstractos. Los lóbulos parietal, temporal y occipital, todos ubicados en la parte posterior de la corteza, integran la información sensorial y la información almacenada en la memoria. El lóbulo frontal o complejo de asociación prefrontal está involucrado en la planificación de acciones y movimientos, así como en el pensamiento abstracto. Globalmente, las áreas de asociación están organizadas como redes distribuidas. Cada red conecta áreas distribuidas en regiones muy espaciadas de la corteza. Distintas redes se colocan adyacentes entre sí, lo que produce una serie compleja de redes entretejidas. La organización específica de las redes de asociaciones se debate con evidencia de interacciones, relaciones jerárquicas y competencia entre redes.
En los seres humanos, las redes de asociación son especialmente importantes para el funcionamiento del lenguaje. En el pasado se teorizó que las habilidades lingüísticas se localizan en el área de Broca en áreas de la circunvolución frontal inferior izquierda, BA44 y BA45, para la expresión del lenguaje y en el área BA22 de Wernicke, para la recepción del lenguaje. Sin embargo, se ha demostrado que los procesos de expresión y recepción del lenguaje ocurren en áreas distintas de las estructuras alrededor del surco lateral, incluido el lóbulo frontal, los ganglios basales, el cerebelo y la protuberancia.
Importancia clínica

Enfermedades neurodegenerativas como la enfermedad de Alzheimer, muestran como marcador, una atrofia de la materia gris de la corteza cerebral.
Otras enfermedades del sistema nervioso central incluyen trastornos neurológicos como la epilepsia, los trastornos del movimiento y diferentes tipos de afasia (dificultades en la expresión o comprensión del habla).
El daño cerebral por enfermedad o trauma puede implicar daño a un lóbulo específico, como en el trastorno del lóbulo frontal, y las funciones asociadas se verán afectadas. La barrera hematoencefálica que sirve para proteger el cerebro de infecciones puede verse comprometida y permitir la entrada de patógenos.
El feto en desarrollo es susceptible a una variedad de factores ambientales que pueden causar defectos de nacimiento y problemas en el desarrollo posterior. El consumo materno de alcohol, por ejemplo, puede causar un trastorno del espectro alcohólico fetal. Otros factores que pueden causar trastornos del neurodesarrollo son los tóxicos como los fármacos y la exposición a radiaciones como los rayos X. Las infecciones también pueden afectar el desarrollo de la corteza. Una infección viral es una de las causas de la lisencefalia, que da como resultado una corteza lisa sin girificación.
Un tipo de electrocorticografía llamado mapeo de estimulación cortical es un procedimiento invasivo que consiste en colocar electrodos directamente en el cerebro expuesto para localizar las funciones de áreas específicas de la corteza. Se utiliza en aplicaciones clínicas y terapéuticas, incluido el mapeo prequirúrgico.
Genes asociados a trastornos corticales
Hay una serie de mutaciones genéticas que pueden causar una amplia variedad de trastornos genéticos de la corteza cerebral, como microcefalia, esquizencefalia y tipos de lisencefalia. Las anomalías cromosómicas también pueden causar una serie de trastornos del desarrollo neurológico, como el síndrome de X frágil y el síndrome de Rett.
MCPH1 codifica para microcefalia, y los trastornos en este y en ASPM se asocian con microcefalia. Las mutaciones en el gen NBS1 que codifica la nibrina pueden causar el síndrome de rotura de Nijmegen, caracterizado por microcefalia.
Las mutaciones en EMX2 y COL4A1 están asociadas con esquizencefalia, una condición caracterizada por la ausencia de gran parte de los hemisferios cerebrales.
Historia
En 1909, Korbinian Brodmann distinguió diferentes áreas de la neocorteza basándose en diferencias citoarquitectónicas y dividió la corteza cerebral en 52 regiones.
Rafael Lorente de Nó, alumno de Santiago Ramón y Cajal identificó más de 40 tipos diferentes de neuronas corticales en función de la distribución de sus dendritas y axones.
Otros animales
La corteza cerebral se deriva del palio, una estructura en capas que se encuentra en el cerebro anterior de todos los vertebrados. La forma básica del palio es una capa cilíndrica que encierra ventrículos llenos de líquido. Alrededor de la circunferencia del cilindro hay cuatro zonas, el palio dorsal, el palio medial, el palio ventral y el palio lateral, que se cree que son homólogos a la neocorteza, el hipocampo, la amígdala y la corteza olfativa, respectivamente.
Hasta hace poco no se había reconocido en los invertebrados ninguna contraparte de la corteza cerebral. Sin embargo, un estudio publicado en la revista Cell en 2010, basado en perfiles de expresión génica, informó fuertes afinidades entre la corteza cerebral y los cuerpos en forma de hongo del gusano de trapo Platynereis dumerilii. Los cuerpos de hongos son estructuras en los cerebros de muchos tipos de gusanos y artrópodos que se sabe que juegan un papel importante en el aprendizaje y la memoria; la evidencia genética indica un origen evolutivo común y, por lo tanto, indica que los orígenes de los primeros precursores de la corteza cerebral se remontan a la era precámbrica.
Imágenes adicionales
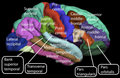
Superficie lateral de la corteza cerebral humana

Superficie medial de la corteza cerebral humana


Contenido relacionado
Jane goodall
Orientación a proteínas
Pigmento fotosintético