
Complejo promotor de la anafase

complejo promotor de la anafase (también llamado ciclosoma o APC/C) es una ubiquitina ligasa E3 que marca las proteínas diana del ciclo celular para degradación por el proteasoma 26S. El APC/C es un gran complejo de proteínas de 11 a 13 subunidades, incluida una subunidad cullin (Apc2) y RING (Apc11) muy parecida a SCF. Otras partes del APC/C tienen funciones desconocidas pero están muy conservadas.
Fue el descubrimiento de APC/C (y SCF) y su papel clave en la regulación del ciclo celular eucariota lo que estableció la importancia de la proteólisis mediada por ubiquitina en la biología celular. La ubiquitinación y la posterior degradación de proteínas por parte del proteasoma, que alguna vez se percibieron como un sistema involucrado exclusivamente en la eliminación de proteínas dañadas de la célula, ahora se perciben como un mecanismo regulador universal para la transducción de señales cuya importancia se acerca a la de la fosforilación de proteínas.
En 2014, el APC/C fue mapeado en 3D con una resolución de menos de un nanómetro, lo que también reveló su estructura secundaria. Este hallazgo podría mejorar la comprensión del cáncer y revelar nuevos sitios de unión para futuros medicamentos contra el cáncer.
Función
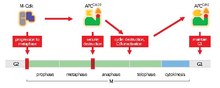
La función principal de APC/C es desencadenar la transición de metafase a anafase marcando proteínas específicas para su degradación. Los tres objetivos principales de degradación por parte del APC/C son la securina y las ciclinas S y M. La securina libera separasa, una proteasa, cuando se degrada. Luego, la separasa desencadena la escisión de la cohesina, el complejo proteico que une las cromátidas hermanas. Durante la metafase, las cromátidas hermanas están unidas por complejos de cohesina intactos. Cuando la securina sufre ubiquitinación por parte de APC/C y libera separasa, que degrada la cohesina, las cromátidas hermanas quedan libres para moverse a polos opuestos para la anafase. El APC/C también se dirige a las ciclinas mitóticas para su degradación, lo que resulta en la inactivación de los complejos M-CDK (quinasa mitótica dependiente de ciclina), promoviendo la salida de la mitosis y la citocinesis.
A diferencia del SCF, las subunidades activadoras controlan el APC/C. Cdc20 y Cdh1 son los dos activadores de particular importancia para el ciclo celular. Estas proteínas dirigen el APC/C a conjuntos específicos de sustratos en diferentes momentos del ciclo celular, impulsándolo así hacia adelante. El APC/C también desempeña un papel integral en el mantenimiento del metabolismo de la cromatina, particularmente en G1 y G0, y desempeña un papel clave en la fosforilación de H3 mediante la destrucción de la quinasa Aurora A.
Los sustratos críticos de APC/C parecen ser la securina y las ciclinas de tipo B. Esto se conserva entre los mamíferos y la levadura. De hecho, las levaduras son viables en ausencia de APC/C si se elimina el requisito de apuntar a estos dos sustratos.
Subunidades
No existe una gran cantidad de investigaciones exhaustivas sobre las subunidades APC/C, que sirven principalmente como adaptadores. Los estudios de las subunidades APC se realizan principalmente en levaduras, y los estudios muestran que la mayoría de las subunidades APC de levadura también están presentes en vertebrados, lo que sugiere conservación entre eucariotas. Se han encontrado once subunidades centrales de APC en vertebrados frente a trece en levaduras. Las subunidades activadoras se unen a APC en distintas etapas del ciclo celular para controlar su actividad de ubiquitinación, a menudo dirigiendo APC a sustratos destinados a la ubiquitinación. Se propone controlar la especificidad de las ligasas APC mediante la incorporación de factores de especificidad en el complejo de ligasa, en lugar de la fosforilación del sustrato. es decir: la subunidad CDC20 permite que APC degrade sustratos como los inhibidores de la anafase (Pdsp1) al comienzo de la anafase; por otro lado, cuando CDC20 se sustituye por el factor de especificidad Hct1, APC degrada un conjunto diferente de sustratos, particularmente las ciclinas de la mitosis al final. anafase. Los activadores CDC20 y Cdh1 son de particular importancia y son las más estudiadas y conocidas de las subunidades APC/C.
El núcleo catalítico de APC/C consta de la subunidad de cullin Apc2 y la subunidad del dominio RING H2 Apc11. Estas dos subunidades catalizan la ubiquitinación de sustratos cuando el dominio C-terminal de Apc2 forma un complejo estrecho con Apc11. RING/APc11 se une al conjugado E2-ubiquitina que cataliza la transferencia de ubiquitina a un sitio activo en E2. Además de la funcionalidad catalítica, otras proteínas centrales de la APC están compuestas por múltiples motivos repetidos con el objetivo principal de proporcionar soporte al andamio molecular. Estos incluyen Apc1, la subunidad más grande que contiene 11 repeticiones en tándem de 35 a 40 secuencias de aminoácidos, y Apc2, que contiene tres repeticiones de cullina de aproximadamente 130 aminoácidos en total. Los motivos principales en las subunidades de APC incluyen motivos tetratricopeptídicos (TPR) y repeticiones de WD40 1. Las regiones C-terminales de CDC20 y Cdh1 tienen un dominio WD40 que se sugiere que forma una plataforma de unión que se une a los sustratos de APC, contribuyendo así a la capacidad de las APC para apuntar a estos. sustratos, aunque se desconoce el mecanismo exacto a través del cual aumentan la actividad de APC. También se sugiere que las variaciones en estos dominios WD40 dan como resultado una especificidad de sustrato variable, lo que se confirma con resultados recientes que sugieren que diferentes sustratos de APC pueden unirse directa y específicamente a Cdc20 y Cdh1/Hct1. En última instancia, las diferencias de especificidad son responsables del momento de la destrucción de varios objetivos de APC durante la mitosis. Con CDC20 apuntando a algunos sustratos principales en la metafase y Cdh1 apuntando a una gama más amplia de sustratos hacia la mitosis tardía y G1.
En particular, 4 subunidades de APC/C de levadura consisten casi en su totalidad en múltiples repeticiones del motivo del residuo tetratricopeptídico (TPR) de 34 aminoácidos. Estas subunidades de TPR, Cdc16, Cdc27, Cdc23 y Apc5, proporcionan principalmente andamiaje y soporte para mediar otras interacciones proteína-proteína. Se ha demostrado que Cdc27 y Cdc23 apoyan la unión de Cdc20 y Cdh1, ya que las mutaciones en residuos clave de estas subunidades condujeron a una mayor disociación de los activadores. Se ha demostrado que Apc10/Doc1 promueve la unión del sustrato mediando sus interacciones con Cdh1 y Cdc20.
En particular, CDC20 (también conocido como p55CDC, Fizzy o Slp1) inactiva CDK1 mediante la ubiquitinación de ciclinas de tipo B. Esto da como resultado la activación de Cdh1 (también conocido como relacionado con Fizzy, Hct1, Ste9 o Srw1), que interactúa con APC durante la mitosis tardía y G1/G0. Cdh1 se inactiva mediante fosforilación durante las fases S, G2 y M temprana. Durante estos puntos del ciclo, no se puede montar.
La evidencia muestra que APC3 y APC7 sirven para reclutar Cdh1 en el complejo promotor de la anafase. Esto respalda aún más que Cdh1 es responsable de mantener la actividad de APC durante G1. Cdh1 no requiere que APC esté fosforilada para unirse; de hecho, la fosforilación de Cdh1 por Cdks evita que se una a APC desde la fase S a M. Con la destrucción de M-Cdk, ahora puede ocurrir la liberación de CDC20 de APC y la unión de Cdh1, lo que permite que la actividad de APC continúe durante la entrada a G1. Mientras que Cdh1 reconoce las ciclinas M y S, lo que permite su destrucción hasta que toda la célula se compromete a proceder a un nuevo ciclo, no reconoce las ciclinas G1/S, y durante la fase G1/S, su actividad de ciclina puede aumentar sin obstáculos y fosforilarse y, por lo tanto, inactivando Cdh1 y por lo tanto APC.
La subunidad Apc15 desempeña un papel importante en la activación de APC/CCdc20 tras la biorientación de las cromátidas hermanas a través de la placa metafásica. Cuando los cinetocoros no están unidos a los husos, los complejos de puntos de control mitóticos (MCC) inhiben la APC. En ausencia de Apc15, los MCC y Cdc20 permanecen bloqueados en el APC/C impidiendo su actividad una vez que se cumplen los requisitos del punto de control del husillo. Apc15 media en la rotación de Cdc20 y MCC para proporcionar información sobre el estado de unión de los cinetocoros.
CDC27/APC3
Se ha identificado que una de las subunidades que exhibe el motivo TPR, CDC27, interactúa con proteínas de punto de control mitótico como Mad2, p55CDC y BUBR1, lo que sugiere que puede estar implicada en el momento de la fase M. La evidencia muestra que CDC27 participa en un complejo ternario con SMAD2/3 y Cdh1, que se crea en respuesta a la señalización de TGFβ. Debido a su interacción con Cdh1 en particular, tiene un papel potencial en la determinación de la afinidad entre APC y sus activadores Cdc20 y Cdh1. Un estudio sugiere que la fosforilación de Cdc27 inducida por TGF-β mejora la interacción entre cdc27 y Cdh1, que participa directamente en la activación de APC. CDC27 puede servir como vehículo a través del cual la señalización de TGFβ puede activar APC. La hiperfosforilación de CDC27 inducida por TGFβ mostró una actividad elevada de APC.
CDC23, CDC16, CDC27
CDC23, otra subunidad de TPR, interactúa con SWM1 y se une a la caja D de CLB2. Con base en ensayos híbridos in vivo y coinmunoprecipitación in vitro, se sugiere que Cdc16p, Cdc23p y Cdc27p (análogos de Sacchyromyces cerevisiae) interactúan y forman un complejo macromolecular. Se sugiere que su tema común, el TPR, media en estas interacciones. En cuanto a Cdc27 y Cdc16 en drosophila, sus funciones se han probado mediante ARN de interferencia (ARNi). Los resultados sugieren que pueden mediar en la actividad de todo el complejo a través de diferentes mecanismos en diferentes sitios. En estudios adicionales de Drosophila, Cdk16 y cdk23 parecen activarse mediante fosforilación por la quinasa 1 similar a Polo (Plk1) y su contraparte de levadura de fisión, parecen unirse particularmente a Cdc23.
Se entiende que el complejo está regulado por los activadores CDC20 y Cdh1 durante la mitosis. Su papel en la degradación de la ciclina B se demuestra mediante una selección de mutantes de Saccharomyces cerevisiae defectuosos para la degradación de la ciclina B, que tenían mutaciones en los genes CDC16 y CDC23. Todos los mutantes de CDC27, CDC23 y CDC 27 provocaron una detención del ciclo celular en la metafase.
Reconocimiento de sustrato
Los sustratos de APC/C tienen secuencias de aminoácidos de reconocimiento que permiten que APC/C los identifique. La secuencia más común se conoce como caja de destrucción o caja D. APC/C reúne una enzima conjugadora de ubiquitina E2 y la caja D en lugar de ser un portador covalente intermedio. La caja D debe tener una versión de la siguiente secuencia de aminoácidos: RXXLXXXXN, donde R es arginina, X es cualquier aminoácido, L es leucina y N es asparagina. El Ken-box es otro motivo de importancia. Su secuencia debe parecerse a la siguiente: KENXXXN, donde K es lisina y E es glutamato. La posición del último aminoácido en la caja Ken es muy variable. Aunque se ha demostrado que las mutaciones en las secuencias inhiben la destrucción de las proteínas "in vivo", todavía hay mucho que aprender sobre cómo las proteínas son atacadas por APC/C.
Una vez unidos a APC/C, Cdc20 y Cdh1 sirven como receptores de caja D y KEN para varios sustratos de APC. Kraft et al. han demostrado que los sustratos' Las cajas D se unen directamente a la región de hélice repetida WD40 altamente conservada en los activadores APC. Es importante señalar que el área conservada de la hélice de Cdh1 es mucho mayor que la de Cdc20, lo que permite que Cdh1 tenga una especificidad de sustrato más amplia, lo que concuerda con el hecho de que APC/CCdh1 también activa APC. -Destrucción mediada por cajas KEN que contienen sustratos. La caja D mejora aún más la degradación de proteínas, ya que los residuos de lisina muy próximos a la caja D sirven como objetivos de ubiquitilación. Se ha descubierto que un residuo de Lys inmediatamente C-terminal de la caja D puede funcionar como aceptor de ubiquitina.
Muchos sustratos de APC contienen cajas D y KEN, y su ubiquitilación por APC/CCdc20 o APC/CCdh1 depende de ambas secuencias, pero algunos sustratos contienen sólo una caja D o una caja KEN, en una o varias copias. Tener dos secuencias de degradación distintas crea un alto nivel de especificidad de sustrato en APC/C, siendo APC/CCdc20 más dependiente de la caja D y APC/CCdh1 más. dependiendo de la caja KEN. Por ejemplo, APC/CCdh1 es capaz de ubiquitilar sustratos que contienen solo cajas KEN como Tome-1 y Sororin.
Aunque Cdc20 y Cdh1 pueden servir como receptores de caja D y KEN, la baja afinidad de estas interacciones coactivador-sustrato sugiere que es poco probable que los coactivadores por sí solos sean suficientes para conferir una unión de sustrato de alta afinidad a la APC. /CCdc20 y APC/CCdh1. En consecuencia, las subunidades centrales de APC/C, como Apc10, también contribuyen a la asociación de sustratos. En construcciones APC/C que carecen de la subunidad Apc10/Doc1, los sustratos como Clb2 no pueden asociarse con APCΔdoc1–Cdh1, mientras que la adición de Doc1 purificado a APCΔdoc1–Cdh1 La construcción restaura la capacidad de unión al sustrato.
Transición de metafase a anafase
A medida que comienza la metafase, el punto de control del huso inhibe el APC/C hasta que todos los cinetocoros hermanos se unen a los polos opuestos del huso mitótico, un proceso conocido como biorientación cromosómica. Cuando todos los cinetocoros están conectados correctamente, el punto de control del husillo se silencia y el APC/C puede activarse. Las M-Cdks fosforilan subunidades en APC/C que promueven la unión a Cdc20. Luego, APC/CCdc20 se dirige a la securina y las ciclinas M (ciclina A y ciclina B) para su degradación. Una vez degradada, se libera separina, se degrada la cohesina y las cromátidas hermanas se preparan para moverse a sus respectivos polos para la anafase.
Es probable que, en las células animales, al menos parte de la activación de APC/CCdc20 ocurra temprano en el ciclo celular (profase o prometafase) según el momento de la degradación de sus sustratos. La ciclina A se degrada temprano en la mitosis, lo que respalda la teoría, pero la ciclina B y la securina no se degradan hasta la metafase. Se desconoce la base molecular del retraso, pero se cree que implica la clave para el momento correcto del inicio de la anafase. En las células animales, el sistema de puntos de control del huso contribuye al retraso si es necesario corregir la biorientación de los cromosomas. Aunque se desconoce cómo el sistema de punto de control del huso inhibe la destrucción de la ciclina B y la securina mientras permite que la ciclina A se degrade. El retraso también puede explicarse por interacciones desconocidas con reguladores, cambios de localización y fosforilación.
Esto inicia un ciclo de retroalimentación negativa. Si bien la activación de APC/CCdc20 requiere M-Cdk, el complejo también es responsable de romper la ciclina para desactivar M-CdK. Esto significa que APC/CCdc20 fomenta su propia desactivación. Es posible que esta retroalimentación negativa sea la columna vertebral de la actividad de Cdk controlada por las oscilaciones de la concentración de ciclina M y S.
Transición de M a G1
Al finalizar la mitosis, es importante que las células (excepto las embrionarias) pasen por un período de crecimiento, conocido como fase G1, para crecer y producir los factores necesarios para el siguiente ciclo celular. La entrada en otra ronda de mitosis se evita inhibiendo la actividad de Cdk. Si bien diferentes procesos son responsables de esta inhibición, uno importante es la activación de APC/C por Cdh1. Esta activación continua previene la acumulación de ciclina que desencadenaría otra ronda de mitosis y, en cambio, impulsa la salida de la mitosis.
Al comienzo del ciclo celular, Cdh1 es fosforilada por M-Cdk, impidiendo que se una a APC/C. Luego, APC/C puede unirse libremente a Cdc20 y marcar el comienzo de la transición de la metafase a la anafase. A medida que M-Cdk se degrada más adelante en la mitosis, Cdc20 se libera y Cdh1 puede unirse a APC/C, manteniéndolo activado durante la transición M/G1. Una diferencia clave a tener en cuenta es que, si bien la unión de Cdc20 a APC/C depende de la fosforilación de APC/C por las Cdks mitóticas, la unión de Cdh1 no lo es. Por lo tanto, a medida que APCCdc20 se inactiva durante la metafase debido a la desfosforilación resultante de las Cdks mitóticas inactivas, Cdh1 puede unirse inmediatamente a APC/C, ocupando el lugar de Cdc20. Cdc20 también es un objetivo de APC/CCdh1, lo que garantiza que APC/CCdc20 esté apagado. APC/CCdh1 luego continúa trabajando en G1 para etiquetar las ciclinas S y M para su destrucción. Sin embargo, las ciclinas G1/S no son sustratos de APC/CCdh1 y, por lo tanto, se acumulan a lo largo de esta fase y fosforilan Cdh1. Al final de G1, se han acumulado suficientes ciclinas G1/S y se ha fosforilado Cdh1 para inactivar APC/C hasta la siguiente metafase.
Una vez en G1, APCCdh1 es responsable de la degradación de varias proteínas que promueven la progresión adecuada del ciclo celular. La geminina es una proteína que se une a Cdt1, lo que impide su unión al complejo de reconocimiento de origen (ORC). APCCdh1 apunta a la geminina para su ubiquitinación en todo G1, manteniendo sus niveles bajos. Esto permite que Cdt1 lleve a cabo su función durante el montaje previo al RC. Cuando APCCdh1 se vuelve inactiva debido a la fosforilación de Cdh1 por las ciclinas G1/S, la actividad de geminina aumenta nuevamente. Además, Dbf4 estimula la actividad de la proteína quinasa (Cdc7) relacionada con el ciclo de división celular 7, que promueve la activación de los orígenes de replicación. Se cree que APCCdh1 apunta a Dbf4 para destruirlo. Esto podría proporcionar una respuesta sobre cómo se activa Cdc7 al comienzo de un nuevo ciclo celular. Su actividad probablemente corresponde a la inactivación de APC/CCdh1 por las ciclinas G/S.
Reglamento adicional
La inactivación deAPC/CCdc20 durante las primeras etapas del ciclo celular se logra parcialmente mediante la proteína Emi1. Los experimentos iniciales han demostrado que la adición de Emi1 a los extractos cíclicos de Xenopus puede prevenir la destrucción de la ciclina A endógena, la ciclina B y la salida mitótica, lo que sugiere que Emi1 es capaz de contrarrestar la actividad de la APC. Además, el agotamiento de Emi1 en las células somáticas conduce a la falta de acumulación de ciclina B. La falta de Emi1 probablemente conduce a una falta de inhibición de la APC que impide que se acumule ciclina B.
A partir de estas primeras observaciones, se ha confirmado que en G2 y en la mitosis temprana, Emi1 se une e inhibe Cdc20 evitando su asociación con sustratos de APC. Cdc20 todavía puede fosforilarse y unirse a APC/C, pero Emi1 unido bloquea la interacción de Cdc20 con los objetivos de APC. La asociación de Emi1 con Cdc20 permite la estabilización de varias ciclinas durante las fases S y G2, pero la eliminación de Emi1 es esencial para la progresión a través de la mitosis. Así, en la profase tardía, Emi1 es fosforilada por la quinasa tipo Polo, Plk. Plk se activa durante la mitosis temprana por la actividad de Cdk1, y su fosforilación del sitio de unión βTrCP BTRC (gen) de Emi1 lo convierte en un objetivo para SCF, lo que lleva a su posterior destrucción en la prometafase. La destrucción de Emi1 conduce a la activación de APC/CCdc20, lo que permite la destrucción de la ciclina A en las primeras etapas de la mitosis. Los niveles de Emi1 comienzan a aumentar nuevamente en G, lo que ayuda a inhibir APC/CCdh1.
La regulación de la actividad de APC/CCdc20 hacia sustratos en metafase como la securina y la ciclina B puede ser el resultado de la localización intracelular. Se plantea la hipótesis de que las proteínas del punto de control del huso que inhiben APC/CCdc20 solo se asocian con un subconjunto de la población de Cdc20 localizada cerca del huso mitótico. De esta manera, la ciclina A se puede degradar, mientras que la ciclina B y la securina se degradan sólo una vez que las cromátidas hermanas han alcanzado la biorientación.