La ciclooxigenasa-2 (COX-2), también conocida como prostaglandina-endoperóxido sintasa 2 (HUGO PTGS2), es una enzima codificada por el gen PTGS2 en humanos. En humanos, es una de las tres ciclooxigenasas. Participa en la conversión del ácido araquidónico en prostaglandina H2, un precursor importante de la prostaciclina, que se expresa en la inflamación.
Función
La PTGS2 (COX-2) convierte el ácido araquidónico (AA) en endoperóxido de prostaglandina H₂. Los PTGS son dianas de los AINE y de los inhibidores específicos de la PTGS2 (COX-2), llamados coxibs. La PTGS-2 es un homodímero de secuencia. Cada monómero de la enzima tiene un sitio activo de peroxidasa y de PTGS (COX). Las enzimas PTGS (COX) catalizan la conversión de AA en prostaglandinas en dos pasos. Primero, se extrae hidrógeno del carbono 13 del ácido araquidónico y, a continuación, la PTGS2 (COX-2) añade dos moléculas de oxígeno, lo que da lugar a PGG2. Segundo, la PGG2 se reduce a PGH2 en el sitio activo de la peroxidasa. La PGH2 sintetizada se convierte en prostaglandinas (PGD2, PGE2, PGF2α), prostaciclina (PGI2) o tromboxano A2 por acción de isomerasas específicas de tejido (Figura 2).Mientras metaboliza el ácido araquidónico principalmente a PGG2, la COX-2 también convierte este ácido graso en pequeñas cantidades de una mezcla racémica de ácidos 15-hidroxiicosatetraenoicos (es decir, 15-HETE) compuesta por aproximadamente un 22 % de estereoisómeros de 15(R)-HETE y aproximadamente un 78 % de 15(S)-HETE, así como una pequeña cantidad de 11(R)-HETE. Los dos estereoisómeros de 15-HETE tienen actividades biológicas intrínsecas, pero, quizás más importante aún, pueden metabolizarse posteriormente a una clase importante de agentes: las lipoxinas. Además, la COX-2 tratada con aspirina metaboliza el ácido araquidónico casi exclusivamente a 15(R)-HETE, producto que puede metabolizarse posteriormente a epilipoxinas. Las lipoxinas y epilipoxinas son potentes agentes antiinflamatorios y pueden contribuir a la actividad general de las dos COX, así como a la aspirina.La COX-2 es inhibida naturalmente por el calcitriol (la forma activa de la vitamina D).
Mecanismo
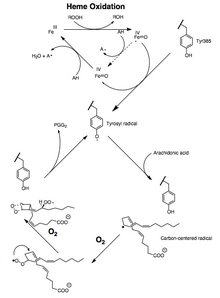
Ácido araquidonico ligado a la enzima PTGS2 (COX-2). Las interacciones polares entre ácido araquidónico (cian) y Ser-530 y los residuos Tyr-385 se muestran con líneas dashed amarillas. El sustrato está estabilizado por interacciones hidrofóbicas. Mecanismo de activación y catalisis de COX. Un hidroperoxido oxida el hemo a un derivado ferryl-oxo que se reduce en el primer paso del ciclo de peroxidase o oxida Tyrosine 385 a un radical de tirosil. El radical de tirosil puede oxidar el hidrógeno de 13-pro(S) de ácido araquidónico para iniciar el ciclo COX.Tanto la actividad de la peroxidasa como la de la PTGS se inactivan durante la catálisis mediante procesos de primer orden basados en mecanismos, lo que significa que la actividad de la peroxidasa PGHS-2 o la de la PTGS se reduce a cero en 1 o 2 minutos, incluso en presencia de suficientes sustratos.La conversión del ácido araquidónico en PGG2 puede mostrarse como una serie de reacciones radicalarias análogas a la autooxidación de ácidos grasos poliinsaturados. El 13-pro(S)-hidrógeno se abstrae y el dioxígeno atrapa el radical pentadienilo en el carbono 11. El radical 11-peroxilo se cicla en el carbono 9 y el radical centrado en el carbono generado en C-8 se cicla en el carbono 12, generando el endoperóxido. El radical alílico generado es atrapado por el dioxígeno en el carbono 15 para formar el radical 15-(S)-peroxilo; este radical luego se reduce a PGG2. Esto está respaldado por la siguiente evidencia: 1) se observa un efecto isotópico cinético significativo para la abstracción del 13-pro(S)-hidrógeno; 2) los radicales centrados en el carbono son atrapados durante la catálisis; 3) Se forman pequeñas cantidades de productos de oxidación debido al atrapamiento de oxígeno de un radical alílico intermedio en las posiciones 13 y 15.Otro mecanismo en el que el 13-pro (S)-hidrógeno se desprotona y el carbanión se oxida a un radical es teóricamente posible. Sin embargo, la oxigenación del ácido 10,10-difluoroaraquidónico a ácido 11-(S)-hidroxieicosa-5,8,12,14-tetraenoico no es consistente con la generación de un intermedio carbaniónico, ya que eliminaría el fluoruro para formar un dieno conjugado. Se ha considerado que la ausencia de productos que contienen endoperóxido derivados del ácido 10,10-difluoroaraquidónico indica la importancia de un carbocatión C-10 en la síntesis de PGG2. Sin embargo, el mecanismo catiónico requiere que la formación de endoperóxido se produzca antes de la eliminación del 13-pro (S)-hidrógeno. Esto no es consistente con los resultados de los experimentos isotópicos de oxigenación del ácido araquidónico.
Estructura
Como se muestra, diferentes ligandos se unen a la subunidad alosterica o catalítica. La subunidad alosterica une un FA no substrato, activando (por ejemplo, ácido palmítico). La subunidad alosterica con ácido graso atado activa la subunidad catalítica disminuyendo el Km para AA.PTGS2 (COX-2) existe como un homodímero, cada monómero con una masa molecular de aproximadamente 70 kDa. Las estructuras terciaria y cuaternaria de las enzimas PTGS1 (COX-1) y PTGS2 (COX-2) son casi idénticas. Cada subunidad tiene tres dominios estructurales diferentes: un dominio corto N-terminal del factor de crecimiento epidérmico (EGF); una porción α-helicoidal de unión a la membrana; y un dominio catalítico C-terminal. Las enzimas PTGS (COX, que puede confundirse con la "citocromo oxidasa") son proteínas de membrana monotópicas; el dominio de unión a la membrana consiste en una serie de hélices α anfipáticas con varios aminoácidos hidrofóbicos expuestos a una monocapa de membrana. PTGS1 (COX-1) y PTGS2 (COX-2) son enzimas bifuncionales que llevan a cabo dos reacciones químicas consecutivas en sitios activos espacialmente distintos, pero acoplados mecánicamente. Tanto los sitios activos de la ciclooxigenasa como de la peroxidasa se encuentran en el dominio catalítico, que representa aproximadamente el 80 % de la proteína. El dominio catalítico es homólogo a las peroxidasas de mamíferos, como la mieloperoxidasa.Se ha descubierto que el PTGS2 (COX-2) humano funciona como un heterodímero conformacional con un monómero catalítico (E-cat) y un monómero alostérico (E-allo). El hemo se une únicamente al sitio de peroxidasa de E-cat, mientras que los sustratos, así como ciertos inhibidores (p. ej., celecoxib), se unen al sitio COX de E-cat. E-cat está regulada por E-allo de forma dependiente del ligando que se une a E-allo. Los ácidos grasos (AG) sustrato y no sustrato, y algunos inhibidores de PTGS (COX) (p. ej., naproxeno), se unen preferentemente al sitio PTGS (COX) de E-allo. El ácido araquidónico puede unirse a E-cat y E-allo, pero la afinidad del AA por E-allo es 25 veces mayor que por Ecat. El ácido palmítico, un estimulador eficaz de huPGHS-2, se une únicamente a E-allo en cocristales de ácido palmítico/PGHS-2 murino. Los AG no sustrato pueden potenciar o atenuar los inhibidores de PTGS (COX) dependiendo del ácido graso y de si el inhibidor se une a E-cat o E-allo. Estudios sugieren que la concentración y composición del conjunto de ácidos grasos libres en el entorno celular donde PGHS-2 funciona, también conocido como tono de AG, es un factor clave que regula la actividad de PGHS-2 y su respuesta a los inhibidores de PTGS (COX).
Significado clínico
NSAID (inhibidor no específico de PTGS2 (COX-2)) flurbiprofen (verde) vinculado a PTGS2 (COX-2). Flurbiprofen se estabiliza mediante interacciones hidrofóbicas y interacciones polares (Tyr-355 y Arg-120). PTGS2 (COX-2) no se expresa en condiciones normales en la mayoría de las células, pero se encuentran niveles elevados durante la inflamación. PTGS1 (COX-1) se expresa constitutivamente en muchos tejidos y es la forma predominante en la mucosa gástrica y en los riñones. La inhibición de PTGS1 (COX-1) reduce la producción basal de PGE2 y PG12 citoprotectoras en el estómago, lo que puede contribuir a la ulceración gástrica. Dado que PTGS2 (COX-2) generalmente se expresa solo en células donde las prostaglandinas están reguladas positivamente (p. ej., durante la inflamación), se sospechaba que los fármacos candidatos que inhiben selectivamente PTGS2 (COX-2) presentaban menos efectos secundarios, pero demostraron aumentar sustancialmente el riesgo de eventos cardiovasculares como infarto de miocardio y accidente cerebrovascular. Dos mecanismos diferentes pueden explicar estos efectos contradictorios. La aspirina en dosis bajas protege contra infartos de miocardio y accidentes cerebrovasculares al impedir que PTGS1 (COX-1) forme una prostaglandina llamada tromboxano A2. Mantiene unidas las plaquetas y promueve la coagulación; su inhibición ayuda a prevenir enfermedades cardíacas. Por otro lado, la PTGS2 (COX-2) es una fuente más importante de prostaglandinas, en particular prostaciclina, presente en el revestimiento de los vasos sanguíneos. La prostaciclina relaja o despega las plaquetas, por lo que los inhibidores selectivos de la COX-2 (coxibs) aumentan el riesgo de eventos cardiovasculares debido a la coagulación.Los antiinflamatorios no esteroideos (AINE) inhiben la producción de prostaglandinas por PTGS1 (COX-1) y PTGS2 (COX-2). Los AINE selectivos para la inhibición de PTGS2 (COX-2) tienen menos probabilidades que los fármacos tradicionales de causar efectos adversos gastrointestinales, pero podrían causar eventos cardiovasculares, como insuficiencia cardíaca, infarto de miocardio y accidente cerebrovascular. Estudios con farmacología y genética humana, roedores manipulados genéticamente, otros modelos animales y ensayos aleatorizados indican que esto se debe a la supresión de las prostaglandinas cardioprotectoras dependientes de PTGS2 (COX-2), en particular la prostaciclina.La expresión de PTGS2 (COX-2) está sobreexpresada en muchos tipos de cáncer. La sobreexpresión de PTGS2 (COX-2), junto con el aumento de la angiogénesis y la expresión de SLC2A1 (GLUT-1), se asocia significativamente con carcinomas de vesícula biliar. Además, la prostaglandina E2 sintasa convierte el producto de PTGS2 (COX-2), PGH2, en PGE2, que a su vez puede estimular la progresión del cáncer. Por consiguiente, la inhibición de PTGS2 (COX-2) podría ser beneficiosa en la prevención y el tratamiento de estos tipos de cáncer.Se detectó la expresión de COX-2 en membranas epirretinianas idiopáticas humanas. El bloqueo de las ciclooxigenasas por lornoxicam en la fase aguda de la inflamación redujo la frecuencia de formación de membranas en un 43 % en el modelo de dispasa de RVP y en un 31 % en el de concanavalina. El lornoxicam no solo normalizó la expresión de ciclooxigenasas en ambos modelos de RVP, sino que también neutralizó los cambios en la retina y el grosor coroideo causados por la inyección de agentes proinflamatorios. Estos hechos subrayan la importancia de las ciclooxigenasas y las prostaglandinas en el desarrollo de RVP.La sobreexpresión del gen PTGS2 también se ha vinculado con múltiples etapas de la reproducción humana. Su presencia se encuentra en la placa coriónica, el epitelio amniótico, los sinciciotrofoblastos, los fibroblastos vellosos, el trofoblasto coriónico y el trofoblasto amniótico, así como en la placa basal de la placenta, las células deciduales y los citotrofoblastos extravellosos. Durante la corioamnionitis/deciduitis, la sobreexpresión de PTGS2 en el amnios y la coriodecidua es uno de los tres efectos limitados de la inflamación uterina. El aumento de la expresión del gen PTGS2 en las membranas fetales se relaciona con la presencia de inflamación, lo que provoca la expresión del gen de las prostaglandinas uterinas y la inmunolocalización de las proteínas de la vía de las prostaglandinas en las células del trofoblasto coriónico y la decidua adyacente. PTGS2 está vinculado con el sistema inflamatorio y se ha observado en leucocitos inflamatorios. Se ha observado una correlación positiva con la expresión de PTGS2 en el amnios durante el parto espontáneo, y se descubrió que su expresión aumenta con la edad gestacional tras el parto, sin observarse cambios en el amnios ni en la coriodecidua durante el parto prematuro o a término. Además, la oxitocina estimula la expresión de PTGS2 en las células del miometrio.Se ha demostrado que los portadores del alelo mutante PTGS2 5939C en la población china Han presentan un mayor riesgo de cáncer gástrico. Además, se encontró una conexión entre la infección por Helicobacter pylori y la presencia del alelo 5939C.Durante un ictus isquémico, la falta de oxígeno y glucosa desencadena una cascada de respuestas inflamatorias que conducen a un aumento de la expresión de COX-2, especialmente en neuronas, células gliales y células endoteliales. Esta sobreexpresión contribuye a la producción de prostaglandinas proinflamatorias como la PGE2, que exacerba el daño neuronal al promover la excitotoxicidad, el estrés oxidativo y la apoptosis. Además, las prostaglandinas derivadas de la COX-2 pueden afectar la integridad de la barrera hematoencefálica (BHE), permitiendo que las células inmunitarias periféricas y los mediadores inflamatorios se infiltren en el cerebro, agravando aún más la lesión cerebral.
Interacciones
Se ha demostrado que PTGS2 interactúa con la caveolina 1.
Historia
La PTGS2 (COX-2) fue descubierta en 1991 por el laboratorio de Daniel Simmons en la Universidad Brigham Young.
Véase también
Ácido Araquidonico
Ciclooxygenasa
Cyclooxygenase 1
NSAID
Descubrimiento y desarrollo de inhibidores selectivos COX-2
Inhibidor selectivo COX-2
Referencias
^ a b cGRCh38: Ensembl release 89: ENSG00073756 – Ensembl, May 2017
^ a b cGRCm38: Ensembl release 89: ENSMUSG00032487 – Ensembl, mayo 2017
^"Human PubMed Referencia:". National Center for Biotechnology Information, U.S. National Library of Medicine.
^"Mouse PubMed Referencia:". National Center for Biotechnology Information, U.S. National Library of Medicine.
^Hla T, Neilson K (agosto de 1992). "Human cyclooxygenase-2 cDNA". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América. 89 (16): 7384–7388. Bibcode:1992PNAS...89.7384H. doi:10.1073/pnas.89.16.7384. PMC 49714. PMID 1380156.
^O'Banion MK (1999). "Cyclooxygenase-2: biología molecular, farmacología y neurobiología". Critical Reviews in Neurobiology. 13 1): 45 –82. doi:10.1615/critrevneurobiol.v13.i1.30. PMID 10223523.
^Mulugeta S, Suzuki T, Hernández NT, Griesser M, Boeglin WE, Schneider C (marzo de 2010). "Identificación y configuración absoluta de ácidos dihidroxi-arachidonicos formados por la oxigenación de 5S-HETE por COX-2 nativo y ácido aspirina". Journal of Lipid Research. 51 3): 575–585. doi:10.1194/jlr.M001719. PMC 2817587. PMID 19752399.
^Serhan CN (2005). "Las lipoxinas y las aspirinas de 15 epi-lipoxinas son los primeros mediadores lípidos de antiinflamación y resolución endógena". Prostaglandinas, Leukotrienes y ácidos grasos esenciales. 73 ()3-4): 141 –162. doi:10.1016/j.plefa.2005.05.002. PMID 16005201.
^Wang Q, He Y, Shen Y, Zhang Q, Chen D, Zuo C, et al. (abril de 2014). "Vitamin D inhibe la expresión COX-2 y la respuesta inflamatoria apuntando a la superfamilia miembro de la tiosterasa 4". El Diario de Química Biológica. 289 (17): 11681–11694. doi:10.1074/jbc.M113.517581. PMC 4002078. PMID 24619416.
^Kassi E, Adamopoulos C, Basdra EK, Papavassiliou AG (diciembre de 2013). "Rolla de vitamina D en aterosclerosis". Circulación. 128 (23): 2517–2531. doi:10.1161/CIRCULATIONAHA.113.002654. PMID 24297817.
^PDB: 3OLT
^Smith WL, Garavito RM, DeWitt DL (diciembre de 1996). "Prostaglandin endoperoxide H synthases (cyclooxygenases)-1 y -2". El Diario de Química Biológica. 271 (52): 33157 –33160. doi:10.1074/jbc.271.52.33157. PMID 8969167.
^Wu G, Wei C, Kulmacz RJ, Osawa Y, Tsai AL (abril de 1999). "Un estudio mecanicista de autoinactivación de la actividad de peroxidasa en prostaglandina H sinthase-1". El Diario de Química Biológica. 274 (14): 9231–9237. doi:10.1074/jbc.274.14.9231. PMID 10092596.
^Callan OH, So OY, Swinney DC (febrero de 1996). "Los factores cinéticos que determinan la afinidad y la selectividad para la lenta inhibición de la prostaglandina humana H sinthasa 1 y 2 por indomethacina y flurbiprofen". El Diario de Química Biológica. 271 (7): 3548–3554. doi:10.1074/jbc.271.7.3548. PMID 8631960.
^Porter NA (1986). "Mecanismos para la autoxidación de lípidos poliinsaturados". Accounts of Chemical Research. 19 (9): 262 –8. doi:10.1021/ar00129a001.
^Mason RP, Kalyanaraman B, Tainer BE, Eling TE (junio de 1980). "Un intermediario radical libre centrado en el carbono en la oxidación de la sintetosis prostaglandina del ácido araquidonico. Estudios de captación y captación de oxígeno". El Diario de Química Biológica. 255 (11): 5019 –5022. doi:10.1016/S0021-9258(19)70741-8. PMID 6246094.
^Hecker M, Ullrich V, Fischer C, Meese CO (noviembre de 1987). "Identificación de nuevos metabolitos de ácido araquidónico formados por prostaglandin H sinthase". European Journal of Biochemistry. 169 1): 113–123. doi:10.1111/j.1432-1033.1987.tb13587.x. PMID 3119336.
^Xiao G, Tsai AL, Palmer G, Boyar WC, Marshall PJ, Kulmacz RJ (febrero de 1997). "Análisis de radicales de tirosil inducidos por hidroperoxido y actividad lipoxigenasa en prostaglandina H sinthase-2 tratada por aspirina". Bioquímica. 36 (7): 1836 –1845. doi:10.1021/bi962476u. PMID 9048568.
^Kwok PY, Muellner FW, Fried J (junio de 1987). "Conversiones enzimáticas de ácido 10,10-difluoroarachidonico con sintosa PGH y lipoxigenasa soja". Journal of the American Chemical Society. 109 (12): 3692 –3698. Bibcode:1987JAChS.109.3692K. doi:10.1021/ja00246a028.
^Dean AM, Dean FM (mayo de 1999). "Carbocaciones en la síntesis de prostaglandinas por la ciclooxigenasa de la sintesis PGH? ¡Una salida radical!". Protein Science. 8 5): 1087 –1098. doi:10.1110/ps.8.5.1087. PMC 2144324. PMID 10338019.
^Hamberg M, Samuelsson B (noviembre de 1967). "En el mecanismo de la biosíntesis de las prostaglandinas E-1 y F-1-alpha". El Diario de Química Biológica. 242 (22): 5336 –5343. doi:10.1016/S0021-9258(18)99433-0. PMID 6070851.
^ a bDong L, Vecchio AJ, Sharma NP, Jurban BJ, Malkowski MG, Smith WL (mayo de 2011). "El ciclooxygenase-2 humano es un homodimer secuencial que funciona como heterodimer conformacional". El Diario de Química Biológica. 286 (21): 19035–19046. doi:10.1074/jbc.M111.231969. PMC 3099718. PMID 21467029.
^Picot D, Loll PJ, Garavito RM (enero de 1994). "La estructura de cristal de rayos X de la proteína de la membrana prostaglandina H2 sinthase-1". Naturaleza. 367 (6460): 243 –249. Bibcode:1994Natur.367..243P. doi:10.1038/367243a0. PMID 8121489. S2CID 4340064.
^ a bKurumbail RG, Kiefer JR, Marnett LJ (diciembre de 2001). "Enzimas ciclooxigenasa: catalisis e inhibición". Opinión actual en Biología Estructural. 11 (6): 752 –760. doi:10.1016/S0959-440X(01)00277-9. PMID 11751058.
^PDB: 3PGH
^Ali A, Wani AB, Malla BA, Poyya J, Dar NJ, Ali F, et al. (Noviembre 2023). "Network Pharmacology Integrated Molecular Docking and Dynamics to Elucidate Saffron Compounds Targeting Human COX-2 Protein". Medicina. 59 (12): 2058. doi:10.3390/medicina59122058. PMC 10744988. PMID 38138161.
^Ruan CH, So SP, Ruan KH (enero de 2011). "El COX-2 inducible domina sobre COX-1 en biosíntesis prostaciclina: mecanismos de riesgo inhibidor COX-2 a enfermedades cardíacas". Ciencias de la vida. 88 ()1 –2): 24 –30. doi:10.1016/j.lfs.2010.10.017. PMC 3046773. PMID 21035466.
^Wang D, Patel VV, Ricciotti E, Zhou R, Levin MD, Gao E, et al. (mayo de 2009). "Cardiomyocyte cyclooxygenase-2 influye en el ritmo cardíaco y la función". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América. 106 (18): 7548 –7552. Bibcode:2009PNAS..106.7548W. doi:10.1073/pnas.0805806106. PMC 2670242. PMID 19376970.
^Legan M (agosto de 2010). "Cyclooxygenase-2, p53 y transportador de glucosa-1 como predictores de malignidad en el desarrollo de carcinomas de vesícula biliar". Bosnian Journal of Basic Medical Sciences. 10 3): 192 –196. doi:10.17305/bjbms.2010.2684. PMC 5504494. PMID 20846124.
^EntrezGene 5743
^Menter DG, Schilsky RL, DuBois RN (marzo de 2010). "Cyclooxygenase-2 y tratamiento del cáncer: entender el riesgo debe valer la recompensa". Clinical Cancer Research. 16 5): 1384–1390. doi:10.1158/1078-0432.CCR-09-0788. PMC 4307592. PMID 20179228.
^Kase S, Saito W, Ohno S, Ishida S (mayo de 2010). "Expresión ciclo-oxigenasa-2 en la membrana epiretina idiopática humana". Retina. 30 5): 719 –723. doi:10.1097/iae.0b013e3181c59698. PMID 19996819. S2CID 205650971.
^Tikhonovich MV, Erdiakov AK, Gavrilova SA (agosto 2018). "La terapia antiinflamatoria no esteroidea suprime el desarrollo de la vitreoretinopatía proliferativa más eficazmente que un esteroide". International Ophthalmology. 38 4): 1365–1378. doi:10.1007/s10792-017-0594-3. PMID 28639085. S2CID 4017540.
^Phillips, Robert J et al. "La expresión del gen de la vía de prostaglandina en la placenta humana, el anión y la choriodecidua se ve afectada de manera diferencial por el trabajo prematuro y el término y por la inflamación uterina". BMC pregnancy and childbirth vol. 14 241. 22 jul. 2014, doi:10.1186/1471-2393-14-241
^Li Y, He W, Liu T, Zhang Q (diciembre de 2010). "Una nueva variante cyclo-oxygenase-2 en la población china de Han está asociada con un mayor riesgo de carcinoma gástrico". Diagnóstico molecular. 14 (6): 351 –355. doi:10.1007/bf03256392. PMID 21275453. S2CID 1229751.
^Wei J, Du K, Cai Q, Ma L, Jiao Z, Tan J, et al. (noviembre de 2014). "Lead induce la expresión COX-2 en células gliales de una manera dependiente de NFAT, AP-1/NFκB-independiente". Toxicology. 325: 67–73. Bibcode:2014Toxgy.325...67W doi:10.1016/j.tox.2014.08.012. PMC 4238429. PMID 25193092.
^Lima IV, Bastos LF, Limborço-Filho M, Fiebich BL, de Oliveira AC (2012). "Rolla de prostaglandinas en enfermedades neuroinflamatorias y neurodegenerativas". Mediadores de Inflamación. 2012 (1): 946813. doi:10.1155/2012/946813. PMC 3385693. PMID 22778499.
^Liou JY, Deng WG, Gilroy DW, Shyue SK, Wu KK (septiembre de 2001). "Colocalización e interacción de ciclooxigenasa-2 con caveolina-1 en fibroblastos humanos". El Diario de Química Biológica. 276 (37): 34975–34982. doi:10.1074/jbc.M105946200. PMID 11432874.
^Xie WL, Chipman JG, Robertson DL, Erikson RL, Simmons DL (abril de 1991). "Expresión de una prostaglandina de codificación de genes resistentes al mitogen está regulada por la rotura de mRNA". Actas de la Academia Nacional de Ciencias de los Estados Unidos de América. 88 (7): 2692 –2696. Código:1991PNAS...88.2692X. doi:10.1073/pnas.88.7.2692. PMC 51304. PMID 1849272.
Más lectura
Richards JA, Petrel TA, Brueggemeier RW (Febrero de 2002). "Patinas de señalización que regulan la aromatasa y cicloxigenas en células mamarias normales y malignas". The Journal of Steroid Biochemistry and Molecular Biology. 80 2): 203 –212. doi:10.1016/S0960-0760(01)00187-X. PMID 11897504. S2CID 12728545.
Wu T, Wu H, Wang J, Wang J (marzo de 2011). "Expresión y localización celular de ciclooxigenas y sintetías prostaglandinas E en el cerebro hemorrágico". Journal of Neuroinflammation. 8: 22. doi:10.1186/1742-2094-8-22. PMC 3062590. PMID 21385433.
Koki AT, Khan NK, Woerner BM, Seibert K, Harmon JL, Dannenberg AJ, et al. (enero de 2002). "Caracterización de la ciclooxigenasa-2 (COX-2) durante la tumorigenesis en cánceres epiteliales humanos: evidencia para la posible utilidad clínica de los inhibidores de COX-2 en cánceres epiteliales". Prostaglandinas, Leukotrienes y ácidos grasos esenciales. 66 1): 13 –18. doi:10.1054/plef.2001.0335. PMID 12051953.
Saukkonen K, Rintahaka J, Sivula A, Buskens CJ, Van Rees BP, Rio MC, et al. (octubre de 2003). "Cyclooxygenasa-2 y carcinogénesis gástrica". APMIS. 111 (10): 915 –925. doi:10.1034/j.1600-0463.2003.1111001.x. PMID 14616542. S2CID 23257867.
Sinicrope FA, Gill S (2004). "Role of cyclooxygenase-2 in colorectal cancer". Cáncer y Metastasis. 23 ()1 –2): 63–75 doi:10.1023/A:1025863029529. PMID 15000150. S2CID 21521040.
Jain S, Khuri FR, Shin DM (2004). "Prevención del cáncer de cabeza y cuello: estado actual y perspectivas futuras". Problemas actuales en el cáncer. 28 5): 265–286. doi:10.1016/j.currproblcancer.2004.05.003. PMID 15375804.
Saba N, Jain S, Khuri F (2004). "Chemoprevención en cáncer de pulmón". Problemas actuales en el cáncer. 28 5): 287 –306. doi:10.1016/j.currproblcancer.2004.05.005. PMID 15375805.
Cardillo I, Spugnini EP, Verdina A, Galati R, Citro G, Baldi A (octubre 2005). "Cox y mesotelioma: una visión general". Histología y Histopatología. 20 4): 1267–1274. doi:10.14670/H-20.1267. PMID 16136507.
Brueggemeier RW, Díaz-Cruz ES (marzo de 2006). "Relación entre aromatasa y ciclooxigenas en cáncer de mama: potencial para nuevos enfoques terapéuticos". Minerva Endocrinologica. 31 1): 13 –26. PMID 16498361.
Fujimura T, Ohta T, Oyama K, Miyashita T, Miwa K (marzo de 2006). "Role of cyclooxygenase-2 in the carcinogenesis of gastrointestinal tract cancers: a review and report of personal experience". World Journal of Gastroenterology. 12 (9): 1336 –1345. doi:10.3748/wjg.v12.i9.1336. PMC 4124307. PMID 16552798.
Bingham S, Beswick PJ, Blum DE, Gray NM, Chessell IP (octubre de 2006). "El papel de la ruta de la citoxigenasa en la nocicepción y el dolor". Seminarios en Celular de Desarrollo Biología. 17 5): 544 –554. doi:10.1016/j.semcdb.2006.09.001. PMID 17071117.
Minghetti L, Pocchiari M (2007). Cyclooxygenase-2, prostaglandin E2, y activación microglial en las enfermedades de los priones. International Review of Neurobiology. Vol. 82. pp. 265–75 doi:10.1016/S0074-7742(07)82014-9. ISBN 978-0-12-373989-6. PMID 17678966.
Enlaces externos
Nextbio Archivado 2019-10-18 en la máquina Wayback
NSAIDs and Cardiovascular Risk Explained, According to Studies from the Perelman School of Medicine
Wolfe MM (diciembre de 2004). "Rofecoxib, Merck y la FDA". El New England Journal of Medicine. 351 (27): 2875–8, respuesta del autor 2875-8. doi:10.1056/NEJM200412303512719. PMID 15625749.
v
t
e
Metabolismo: metabolismo de los lípidos – enzimas del metabolismo eicosanoide