
Ciclo de la urea
El ciclo de la urea (también conocido como ciclo de la ornitina) es un ciclo de reacciones bioquímicas que produce urea (NH2)2CO del amoníaco (NH3). Los animales que utilizan este ciclo, principalmente anfibios y mamíferos, se denominan ureotélicos.
El ciclo de la urea convierte el amoníaco altamente tóxico en urea para su excreción. Este ciclo fue el primer ciclo metabólico descubierto (Hans Krebs y Kurt Henseleit, 1932), cinco años antes del descubrimiento del ciclo TCA. Este ciclo fue descrito con más detalle más adelante por Ratner y Cohen. El ciclo de la urea tiene lugar principalmente en el hígado y, en menor medida, en los riñones.
Función
El catabolismo de aminoácidos da como resultado amoníaco de desecho. Todos los animales necesitan una forma de excretar este producto. La mayoría de los organismos acuáticos, u organismos amonotélicos, excretan amoníaco sin convertirlo. Los organismos que no pueden eliminar de manera fácil y segura el nitrógeno como amoníaco, lo convierten en una sustancia menos tóxica, como la urea, a través del ciclo de la urea, que ocurre principalmente en el hígado. La urea producida por el hígado luego se libera en el torrente sanguíneo, donde viaja a los riñones y finalmente se excreta en la orina. El ciclo de la urea es fundamental para estos organismos, ya que si no se elimina el nitrógeno o el amoníaco del organismo puede ser muy perjudicial. En especies que incluyen aves y la mayoría de los insectos, el amoníaco se convierte en ácido úrico o su sal de urato, que se excreta en forma sólida. Además, el ciclo de la urea consume dióxido de carbono residual ácido al combinarlo con el amoníaco básico, lo que ayuda a mantener un pH neutro.
Reacciones
Todo el proceso convierte dos grupos amino, uno de NH+
4 y uno de aspartato, y un átomo de carbono de HCO−
3, al producto de excreción relativamente no tóxico urea. Esto ocurre a costa de cuatro "alta energía" enlaces fosfato (3 ATP hidrolizados a 2 ADP y uno AMP). La conversión de amoníaco a urea ocurre en cinco pasos principales. El primero es necesario para que el amoniaco entre en el ciclo y los cuatro siguientes forman parte del ciclo mismo. Para entrar en el ciclo, el amoniaco se convierte en fosfato de carbamoilo. El ciclo de la urea consta de cuatro reacciones enzimáticas: una mitocondrial y tres citosólicas. Esto utiliza 6 enzimas.
| Paso | Reactantes | Productos | Catalyzed by | Ubicación |
|---|---|---|---|---|
| 1 | NH3 + HCO− 3 + 2ATP | carbamoil fosfato + 2ADP + Pi | CPS1 | mitocondria |
| 2 | carbamoil fosfato + ornitina | citrulline + Pi | OTC, zinc, biotina | mitocondria |
| 3 | citrulline + aspartate + ATP | argininosuccinate + AMP + PPi | ASS | cytosol |
| 4 | argininosuccinate | arginina + fumarate | ASL | cytosol |
| 5 | arginina + H2O | ornitina + urea | ARG1, manganeso | cytosol |
- Las reacciones del ciclo de la urea
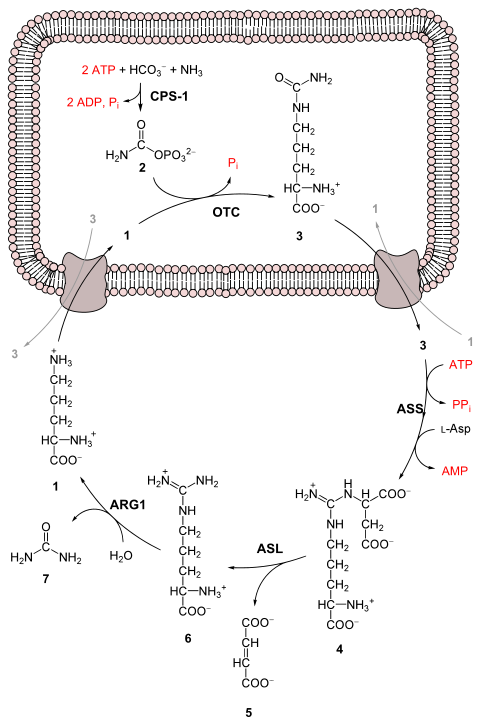
1 L- ornithine
2 fosfato de carbamoilo
3 L-citrulline
4 argininosuccinate
5 fumarate
6 L- Arginina
7 urea
L-Asp L- Aspartate
CPS-1 carbamoil fosfato sintetase I
OTC Ornithine transcarbamoylase
ASS argininosuccinate synthetase
lisa de argininosuccino ASL
ARG1 arginase 1

Primera reacción: entrar en el ciclo de la urea
Antes de que comience el ciclo de la urea, el amoníaco se convierte en fosfato de carbamoilo. La reacción es catalizada por carbamoil fosfato sintetasa I y requiere el uso de dos moléculas de ATP. El fosfato de carbamoilo luego ingresa al ciclo de la urea.
Pasos del ciclo de la urea
- Carbamoyl phosphate se convierte en citrulline. Con catalisis por ornitina transcarbamylase, el grupo de fosfato de carbamoilo se dona a ornitina y libera un grupo de fosfato.
- Una reacción de condensación ocurre entre el grupo amino de aspartato y el grupo de carbonilo de citrullina para formar argininosuccinate. Esta reacción es dependiente de ATP y es catalizada por sintetasa argininosuccinate.
- Argininosuccinate sufre escote por argininosuccinase para formar arginina y fumarato.
- La arginana es cleaveda por la arginasa para formar urea y ornitina. La ornitina es transportada de vuelta a la mitocondria para comenzar el ciclo de urea de nuevo.
Ecuación de reacción global
En la primera reacción, NH+
4 + HCO−
3 es equivalente a NH3 + CO2 + H2O.
Por lo tanto, la ecuación general del ciclo de la urea es:
- NH3 + CO2 + aspartate + 3 ATP + 3 H2O → urea + fumarate + 2 ADP + 2 Pi + AMP + PPi + H2O
Dado que el fumarato se obtiene eliminando el NH3 del aspartato (mediante las reacciones 3 y 4), y PPi + H2 O → 2 Pi, la ecuación se puede simplificar de la siguiente manera:
- 2 NH3 + CO2 + 3 ATP + 3 H2O → urea + 2 ADP + 4 Pi + AMP
Tenga en cuenta que las reacciones relacionadas con el ciclo de la urea también provocan la producción de 2 NADH, por lo que la reacción general libera un poco más de energía de la que consume. El NADH se produce de dos formas:
- Una molécula NADH es producida por la enzima glutamato deshidrogenasa en la conversión de glutamato a amonio y α-ketoglutaato. Glutamate es el portador no tóxico de los grupos de amina. Esto proporciona el ión de amonio utilizado en la síntesis inicial del fosfato de carbamoilo.
- La fumarada liberada en el citosol es hidratada a malate por citosolic fumarase. Este malete es entonces oxidado a oxaloacetate por malete citosolico deshidrogenasa, generando una NADH reducida en el citosol. Oxaloacetate es uno de los ácidos ceto preferidos por las transaminasas, y así será reciclado para aspartar, manteniendo el flujo de nitrógeno en el ciclo de urea.
Podemos resumir esto combinando las reacciones:
- CO2 + glutamato + aspartato + 3 ATP + 2 NAD++ 3 H2O → urea + α-ketoglutarate + oxaloacetate + 2 ADP + 2 Pi + AMP + PPi + 2 NADH
Los dos NADH producidos pueden proporcionar energía para la formación de 5 ATP (el NADH citosólico proporciona 2,5 ATP con la lanzadera malato-aspartato en las células hepáticas humanas), una producción neta de dos enlaces fosfato de alta energía para el ciclo de la urea. Sin embargo, si la gluconeogénesis está en marcha en el citosol, el último equivalente reductor se usa para impulsar la reversión del paso GAPDH en lugar de generar ATP.
El destino del oxaloacetato es producir aspartato a través de la transaminación o convertirse en fosfoenolpiruvato, que es un sustrato para la gluconeogénesis.
Productos del ciclo de la urea
Como se mencionó anteriormente, muchos vertebrados usan el ciclo de la urea para crear urea a partir de amonio para que el amonio no dañe el cuerpo. Aunque esto es útil, existen otros efectos del ciclo de la urea. Por ejemplo: consumo de dos ATP, producción de urea, generación de H+, combinación de HCO−3 y NH+ 4 a formas donde se puede regenerar, y finalmente el consumo de NH+4.
Regulación
Ácido N-acetilglutámico
La síntesis de carbamoil fosfato y el ciclo de la urea dependen de la presencia de ácido N-acetilglutámico (NAcGlu), que activa alostéricamente CPS1. NAcGlu es un activador obligado de la carbamoil fosfato sintetasa. La síntesis de NAcGlu por la N-acetilglutamato sintasa (NAGS) es estimulada tanto por Arg, estimulador alostérico de NAGS, como por Glu, un producto de las reacciones de transaminación y uno de los sustratos de NAGS, ambos elevados cuando los aminoácidos libres son elevados. Entonces, Glu no solo es un sustrato para NAGS, sino que también sirve como activador para el ciclo de la urea.
Concentraciones de sustrato
Las enzimas restantes del ciclo están controladas por las concentraciones de sus sustratos. Por lo tanto, las deficiencias heredadas en las enzimas del ciclo distintas de ARG1 no dan como resultado disminuciones significativas en la producción de urea (si falta por completo alguna enzima del ciclo, la muerte ocurre poco después del nacimiento). Más bien, el sustrato de la enzima deficiente se acumula, aumentando la velocidad de la reacción deficiente a la normalidad.
Sin embargo, la acumulación anómala del sustrato tiene un costo. Las concentraciones de sustrato se elevan durante todo el ciclo hasta NH+
4, lo que provoca hiperamonemia (elevado [NH+
4]P).
Aunque la causa raíz de NH+
4 la toxicidad no se entiende completamente, un alto [NH+
4] ejerce una enorme presión sobre el NH+
4 -sistema de limpieza, especialmente en el cerebro (los síntomas de las deficiencias de las enzimas del ciclo de la urea incluyen discapacidad intelectual y letargo). Este sistema de compensación involucra a GLUD1 y GLUL, que disminuyen las reservas de 2-oxoglutarato (2OG) y Glu. El cerebro es más sensible al agotamiento de estas reservas. El agotamiento de 2OG disminuye la tasa de TCAC, mientras que Glu es tanto un neurotransmisor como un precursor de GABA, otro neurotransmisor. [1](p.734)
Vínculo con el ciclo del ácido cítrico
El ciclo de la urea y el ciclo del ácido cítrico son ciclos independientes pero están vinculados. Uno de los átomos de nitrógeno en el ciclo de la urea se obtiene de la transaminación de oxalacetato a aspartato. El fumarato que se produce en el paso tres también es un intermediario en el ciclo del ácido cítrico y se devuelve a ese ciclo.
Trastornos del ciclo de la urea
Los trastornos del ciclo de la urea son raros y afectan aproximadamente a una de cada 35 000 personas en los Estados Unidos. Pueden ocurrir defectos genéticos en las enzimas involucradas en el ciclo, que generalmente se manifiestan a los pocos días después del nacimiento. El niño recién nacido normalmente experimentará episodios variados de vómitos y períodos de letargo. En última instancia, el bebé puede entrar en coma y desarrollar daño cerebral. Los recién nacidos con UCD tienen un riesgo mucho mayor de complicaciones o muerte debido a pruebas de detección inoportunas y casos mal diagnosticados. El diagnóstico erróneo más común es la sepsis neonatal. Los signos de UCD pueden estar presentes dentro de los primeros 2 a 3 días de vida, pero el método actual para obtener la confirmación de los resultados de las pruebas puede llevar demasiado tiempo. Esto puede causar potencialmente complicaciones como el coma o la muerte.
Los trastornos del ciclo de la urea también se pueden diagnosticar en adultos y los síntomas pueden incluir episodios de delirio, letargo y síntomas similares a los de un derrame cerebral. Además de estos síntomas, si el ciclo de la urea comienza a funcionar mal en el hígado, el paciente puede desarrollar cirrosis. Esto también puede provocar sarcopenia (pérdida de masa muscular). Las mutaciones conducen a deficiencias de las diversas enzimas y transportadores involucrados en el ciclo de la urea y causan trastornos del ciclo de la urea. Si las personas con un defecto en cualquiera de las seis enzimas utilizadas en el ciclo ingieren más aminoácidos de los necesarios para los requisitos mínimos diarios, entonces el amoníaco que se produce no podrá convertirse en urea. Estos individuos pueden experimentar hiperamonemia o la acumulación de un ciclo intermedio.
Trastornos individuales
- N-Acetylglutamate synthase (NAGS) deficiencia
- Carbamoil fosfato sintetasa (CPS) deficiencia
- Deficiencia de Ornithine transcarbamoylase (OTC)
- Citrullinemia tipo I (Deficiencia de la sintesis de ácido argininosuccinico)
- Argininosuccinic aciduria (Deficiencia de la lisa de ácido argininosuccinico)
- Argininemia (Deficiencia de la arginasa)
- Hyperornithinemia, hiperammonemia, homocitrullinuria (HHH) síndrome (Deficiencia del transportador de ornitina mitocondrial)
Todos los defectos del ciclo de la urea, excepto la deficiencia de OTC, se heredan de forma autosómica recesiva. La deficiencia de OTC se hereda como un trastorno recesivo ligado al cromosoma X, aunque algunas mujeres pueden mostrar síntomas. La mayoría de los trastornos del ciclo de la urea están asociados con hiperamonemia, sin embargo, la argininemia y algunas formas de aciduria argininosuccínico no se presentan con amoníaco elevado.
Imágenes adicionales

Ciclo de Urea.

Ciclo de Urea coloreado.


Contenido relacionado
Dorothy hodgkin
Tirosina
Kary mullis