
Ciclo de Calvin

El ciclo de Calvin, reacciones independientes de la luz, fase biosintética, reacciones oscuras o ciclo de reducción de carbono fotosintético (PCR) de la fotosíntesis son las reacciones químicas que convierten el dióxido de carbono y los compuestos portadores de hidrógeno en glucosa. El ciclo de Calvin está presente en todos los eucariotas fotosintéticos y también en muchas bacterias fotosintéticas. En las plantas, estas reacciones ocurren en el estroma, la región llena de líquido de un cloroplasto fuera de las membranas tilacoides. Estas reacciones toman los productos (ATP y NADPH) de reacciones dependientes de la luz y realizan otros procesos químicos sobre ellos. El ciclo de Calvin utiliza la energía química del ATP y el poder reductor del NADPH de las reacciones dependientes de la luz para producir azúcares para uso de la planta. Estos sustratos se utilizan en una serie de reacciones de reducción-oxidación para producir azúcares en un proceso gradual; no hay una reacción directa que convierta varias moléculas de CO 2a un azúcar. Hay tres fases en las reacciones independientes de la luz, denominadas colectivamente ciclo de Calvin: carboxilación, reacciones de reducción y regeneración de ribulosa 1,5-bisfosfato (RuBP).
Aunque se llama la "reacción oscura", el ciclo de Calvin en realidad no ocurre en la oscuridad o durante la noche. Esto se debe a que el proceso requiere NADPH, que es de corta duración y proviene de las reacciones dependientes de la luz. En la oscuridad, las plantas liberan sacarosa en el floema desde sus reservas de almidón para proporcionar energía a la planta. Por lo tanto, el ciclo de Calvin ocurre cuando la luz está disponible independientemente del tipo de fotosíntesis (fijación de carbono C3, fijación de carbono C4 y metabolismo del ácido crasuláceo (CAM)); Las plantas CAM almacenan ácido málico en sus vacuolas todas las noches y lo liberan durante el día para que este proceso funcione.
Acoplamiento a otras rutas metabólicas
Las reacciones del ciclo de Calvin están estrechamente acopladas a la cadena de transporte de electrones de los tilacoides, ya que la energía necesaria para reducir el dióxido de carbono la proporciona el NADPH producido durante las reacciones dependientes de la luz. El proceso de fotorrespiración, también conocido como ciclo C2, también está acoplado al ciclo de Calvin, ya que resulta de una reacción alternativa de la enzima RuBisCO, y su subproducto final es otra molécula de gliceraldehído-3-P.
Ciclo de Calvin

El ciclo de Calvin, ciclo de Calvin-Benson-Bassham (CBB), ciclo de pentosa fosfato reductor (ciclo RPP) o ciclo C3 es una serie de reacciones redox bioquímicas que tienen lugar en el estroma del cloroplasto en organismos fotosintéticos. El ciclo fue descubierto en 1950 por Melvin Calvin, James Bassham y Andrew Benson en la Universidad de California, Berkeley, utilizando el isótopo radiactivo carbono-14.
La fotosíntesis ocurre en dos etapas en una célula. En la primera etapa, las reacciones dependientes de la luz capturan la energía de la luz y la utilizan para producir la molécula de almacenamiento de energía ATP y el transportador de hidrógeno de energía moderada NADPH. El ciclo de Calvin utiliza estos compuestos para convertir el dióxido de carbono y el agua en compuestos orgánicos que pueden ser utilizados por el organismo (y por los animales que se alimentan de él). A este conjunto de reacciones también se le llama fijación de carbono. La enzima clave del ciclo se llama RuBisCO. En las siguientes ecuaciones bioquímicas, las especies químicas (fosfatos y ácidos carboxílicos) existen en equilibrio entre sus diversos estados ionizados según lo rige el pH.
Las enzimas del ciclo de Calvin son funcionalmente equivalentes a la mayoría de las enzimas utilizadas en otras vías metabólicas, como la gluconeogénesis y la vía de las pentosas fosfato, pero las enzimas del ciclo de Calvin se encuentran en el estroma del cloroplasto en lugar del citosol celular, separando las reacciones. Se activan con la luz (por lo que el nombre de "reacción oscura" es engañoso), y también por productos de la reacción dependiente de la luz. Estas funciones reguladoras evitan que el ciclo de Calvin se respire a dióxido de carbono. Se desperdiciaría energía (en forma de ATP) al llevar a cabo estas reacciones cuando no tienen productividad neta.
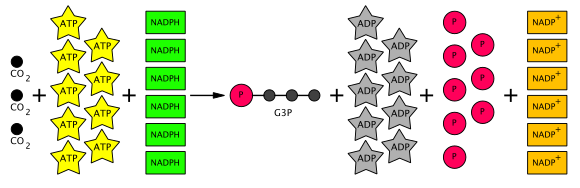
La suma de reacciones en el ciclo de Calvin es la siguiente:3 CO2+ 6 NADPH + 6 H + 9 ATP + 5 H2O → gliceraldehído-3-fosfato (G3P) + 6 NADP + 9 ADP + 8 P i (P i = fosfato inorgánico)
Los azúcares de hexosa (seis carbonos) no son productos del ciclo de Calvin. Aunque muchos textos enumeran un producto de la fotosíntesis como C6H12O6, esto es principalmente por conveniencia para igualar la ecuación de la respiración aeróbica, donde los azúcares de seis carbonos se oxidan en las mitocondrias. Los productos de carbohidratos del ciclo de Calvin son moléculas de fosfato de azúcar de tres carbonos, o "fosfatos de triosa", a saber, gliceraldehído-3-fosfato (G3P).
Pasos
En la primera etapa del ciclo de Calvin, una molécula de CO 2 se incorpora a una de dos moléculas de tres carbonos (gliceraldehído 3-fosfato o G3P), donde consume dos moléculas de ATP y dos moléculas de NADPH, que se habían producido en la etapa dependiente de la luz. Los tres pasos involucrados son:
- La enzima RuBisCO cataliza la carboxilación de ribulosa-1,5-bisfosfato, RuBP, un compuesto de 5 carbonos, por dióxido de carbono (un total de 6 carbonos) en una reacción de dos pasos. El producto del primer paso es un complejo enediol-enzima que puede capturar CO2o O2. Por tanto, el complejo enediol-enzima es la verdadera carboxilasa/oxigenasa. el CO2que es capturado por enediol en el segundo paso produce un compuesto inestable de seis carbonos llamado 2-carboxi 3-ceto 1,5-bifosforribotol (CKABP) (o 3-ceto-2-carboxiarabinitol 1,5-bisfosfato) que inmediatamente se divide en 2 moléculas de 3-fosfoglicerato (también escrito como ácido 3-fosfoglicérico, PGA, 3PGA o 3-PGA), un compuesto de 3 carbonos.
- La enzima fosfoglicerato quinasa cataliza la fosforilación de 3-PGA por ATP (que se produce en la etapa dependiente de la luz). Los productos son 1,3-bisfosfoglicerato (glicerato-1,3-bisfosfato) y ADP. (Sin embargo, tenga en cuenta que se producen dos 3-PGA por cada CO2que entra en el ciclo, por lo que este paso utiliza dos ATP por CO2reparado.)
- La enzima gliceraldehído 3-fosfato deshidrogenasa cataliza la reducción de 1,3BPGA por NADPH (que es otro producto de la etapa dependiente de la luz). Se produce gliceraldehído 3-fosfato (también llamado G3P, GP, TP, PGAL, GAP) y el propio NADPH se oxida y se convierte en NADP. Nuevamente, se utilizan dos NADPH por CO2reparado.
La siguiente etapa en el ciclo de Calvin es regenerar RuBP. Cinco moléculas de G3P producen tres moléculas de RuBP, consumiendo tres moléculas de ATP. Dado que cada molécula de CO 2 produce dos moléculas de G3P, tres moléculas de CO 2 producen seis moléculas de G3P, de las cuales cinco se utilizan para regenerar RuBP, dejando una ganancia neta de una molécula de G3P por cada tres moléculas de CO 2 (como cabría esperar del número de átomos de carbono involucrados).
La etapa de regeneración se puede dividir en una serie de pasos.
- La triosa fosfato isomerasa convierte todo el G3P de forma reversible en fosfato de dihidroxiacetona (DHAP), también una molécula de 3 carbonos.
- La aldolasa y la fructosa-1,6-bisfosfatasa convierten un G3P y un DHAP en fructosa 6-fosfato (6C). Un ion de fosfato se pierde en la solución.
- Luego fijación de otro CO2genera dos G3P más.
- F6P tiene dos carbonos eliminados por la transcetolasa, dando eritrosa-4-fosfato (E4P). Los dos carbonos de la transcetolasa se agregan a un G3P, dando la cetosa xilulosa-5-fosfato (Xu5P).
- E4P y un DHAP (formado a partir de uno de los G3P del segundo CO2fijación) se convierten en sedoheptulosa-1,7-bisfosfato (7C) por la enzima aldolasa.
- La sedoheptulosa-1,7-bisfosfatasa (una de las tres únicas enzimas del ciclo de Calvin que son exclusivas de las plantas) escinde la sedoheptulosa-1,7-bisfosfato en sedoheptulosa-7-fosfato, liberando un ión de fosfato inorgánico en la solución.
- Fijación de un tercer CO2genera dos G3P más. La cetosa S7P tiene dos carbonos eliminados por la transcetolasa, dando ribosa-5-fosfato (R5P), y los dos carbonos que quedan en la transcetolasa se transfieren a uno de los G3P, dando otro Xu5P. Esto deja un G3P como producto de la fijación de 3 CO2, con generación de tres pentosas que se pueden convertir a Ru5P.
- R5P se convierte en ribulosa-5-fosfato (Ru5P, RuP) por la fospentosa isomerasa. Xu5P se convierte en RuP por fospentosa epimerasa.
- Finalmente, la fosforibuloquinasa (otra enzima de la vía exclusiva de la planta) fosforila RuP en RuBP, ribulosa-1,5-bisfosfato, completando el ciclo de Calvin. Esto requiere la entrada de un ATP.
Por lo tanto, de seis G3P producidos, cinco se utilizan para hacer tres moléculas de RuBP (5C) (que suman 15 carbonos), con solo un G3P disponible para su posterior conversión en hexosa. Esto requiere nueve moléculas de ATP y seis moléculas de NADPH por tres CO2moléculas. La ecuación del ciclo total de Calvin se muestra esquemáticamente a continuación.
RuBisCO también reacciona competitivamente con O2en lugar de CO2en fotorrespiración. La tasa de fotorrespiración es mayor a altas temperaturas. La fotorrespiración convierte a RuBP en 3-PGA y 2-fosfoglicolato, una molécula de 2 carbonos que se puede convertir a través de glicolato y glioxalato en glicina. A través del sistema de escisión de glicina y tetrahidrofolato, dos glicinas se convierten en serina más CO2. La serina se puede volver a convertir en 3-fosfoglicerato. Por lo tanto, solo 3 de 4 carbonos de dos fosfoglicolatos se pueden volver a convertir en 3-PGA. Se puede observar que la fotorrespiración tiene consecuencias muy negativas para la planta, ya que más que fijar el CO2, este proceso conduce a la pérdida de CO2. La fijación de carbono C4 evolucionó para eludir la fotorrespiración, pero solo puede ocurrir en ciertas plantas nativas de climas muy cálidos o tropicales, como el maíz, por ejemplo.
Productos
Los productos inmediatos de una vuelta del ciclo de Calvin son 2 moléculas de gliceraldehído-3-fosfato (G3P), 3 ADP y 2 NADP. (ADP y NADPno son realmente "productos". Se regeneran y luego se utilizan de nuevo en las reacciones dependientes de la luz). Cada molécula de G3P está compuesta por 3 carbonos. Para que el ciclo de Calvin continúe, se debe regenerar RuBP (ribulosa 1,5-bisfosfato). Entonces, 5 de los 6 carbonos de las 2 moléculas G3P se usan para este propósito. Por lo tanto, solo se produce 1 carbono neto para jugar en cada turno. Para crear 1 G3P excedente se requieren 3 carbonos y, por lo tanto, 3 vueltas del ciclo de Calvin. Para hacer una molécula de glucosa (que se puede crear a partir de 2 moléculas de G3P) se necesitarían 6 vueltas del ciclo de Calvin. El excedente de G3P también se puede usar para formar otros carbohidratos como almidón, sacarosa y celulosa, según lo que necesite la planta.
Regulación dependiente de la luz

Estas reacciones no ocurren en la oscuridad o en la noche. Hay una regulación dependiente de la luz de las enzimas del ciclo, ya que el tercer paso requiere NADPH.
Hay dos sistemas de regulación en funcionamiento cuando el ciclo debe encenderse o apagarse: el sistema de activación de tiorredoxina/ferredoxina, que activa algunas de las enzimas del ciclo; y la activación de la enzima RuBisCo, activa en el ciclo de Calvin, que implica su propia activasa.
El sistema tiorredoxina/ferredoxina activa las enzimas gliceraldehído-3-P deshidrogenasa, gliceraldehído-3-P fosfatasa, fructosa-1,6-bisfosfatasa, sedoheptulosa-1,7-bisfosfatasa y ribulosa-5-fosfatasa quinasa, que son puntos clave del proceso Esto sucede cuando hay luz disponible, ya que la proteína ferredoxina se reduce en el complejo del fotosistema I de la cadena de electrones tilacoides cuando los electrones circulan a través de él.Ferredoxin luego se une a la proteína tiorredoxina y la reduce, lo que activa las enzimas del ciclo al romper un enlace de cistina que se encuentra en todas estas enzimas. Este es un proceso dinámico ya que el mismo enlace se forma nuevamente por otras proteínas que desactivan las enzimas. Las implicaciones de este proceso son que la mayoría de las enzimas permanecen activadas durante el día y se desactivan en la oscuridad cuando no hay más ferredoxina reducida disponible.
La enzima RuBisCo tiene su propio proceso de activación más complejo. Requiere que un aminoácido de lisina específico sea carbamilado para activar la enzima. Esta lisina se une a RuBP y conduce a un estado no funcional si se deja sin carbamilar. Una enzima activasa específica, llamada RuBisCo activasa, ayuda a este proceso de carbamilación al eliminar un protón de la lisina y hacer posible la unión de la molécula de dióxido de carbono. Incluso entonces, la enzima RuBisCo aún no es funcional, ya que necesita un ion de magnesio unido a la lisina para funcionar. Este ion de magnesio se libera de la luz del tilacoides cuando el pH interno cae debido al bombeo activo de protones del flujo de electrones. La propia RuBisCo activasa se activa por el aumento de las concentraciones de ATP en el estroma causado por su fosforilación.
Contenido relacionado
Grupo metilo
Grupo del platino
Hermann Kolbe