
ADN ligasa
La ligasa de ADN es un tipo específico de enzima, una ligasa (EC 6.5.1.1) que facilita la unión de hebras de ADN al catalizar la formación de un enlace fosfodiéster. Desempeña un papel en la reparación de roturas de una sola cadena en el ADN dúplex en los organismos vivos, pero algunas formas (como la ADN ligasa IV) pueden reparar específicamente roturas de doble cadena (es decir, una rotura en ambas cadenas complementarias de ADN). Las roturas de una sola cadena son reparadas por la ADN ligasa utilizando la cadena complementaria de la doble hélice como plantilla, y la ADN ligasa crea el enlace fosfodiéster final para reparar completamente el ADN.
La ligasa de ADN se utiliza tanto en la reparación como en la replicación del ADN (consulte Ligasas de mamíferos). Además, la ADN ligasa tiene un amplio uso en laboratorios de biología molecular para experimentos de ADN recombinante (ver Aplicaciones de investigación). La ligasa de ADN purificada se usa en la clonación de genes para unir moléculas de ADN para formar ADN recombinante.
Mecanismo enzimático
El mecanismo de la ligasa de ADN es formar dos enlaces fosfodiéster covalentes entre los extremos hidroxilo 3' de un nucleótido ("aceptor"), con el extremo fosfato 5' de otro ("donante"). Se consumen dos moléculas de ATP por cada enlace fosfodiéster formado. Se requiere AMP para la reacción de la ligasa, que se desarrolla en cuatro pasos:
- Reorganización del sitio de actividad, como muescas en segmentos de ADN o fragmentos de Okazaki, etc.
- Adenilación (adición de AMP) de un residuo de lisina en el centro activo de la enzima, se libera pirofosfato;
- Transferencia del AMP al fosfato 5' del llamado donante, formación de un enlace pirofosfato;
- Formación de un enlace fosfodiéster entre el fosfato 5' del donante y el hidroxilo 3' del aceptor.
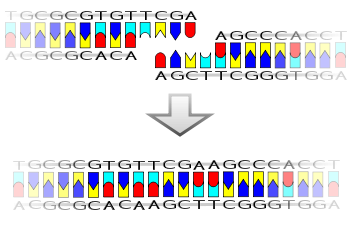 Un ejemplo pictórico de cómo funciona una ligasa (con extremos pegajosos)
Un ejemplo pictórico de cómo funciona una ligasa (con extremos pegajosos)
La ligasa también funcionará con extremos romos, aunque se requieren concentraciones de enzima más altas y diferentes condiciones de reacción.
Tipos
E. coli
La ADN ligasa de E. coli está codificada por el gen lig. La ligasa de ADN en E. coli, así como en la mayoría de los procariotas, utiliza la energía obtenida al escindir el dinucleótido de nicotinamida y adenina (NAD) para crear el enlace fosfodiéster. No liga el ADN de extremos romos, excepto en condiciones de apiñamiento molecular con polietilenglicol, y no puede unir eficientemente el ARN al ADN.
La actividad de la ADN ligasa de E. coli se puede mejorar con la ADN polimerasa en las concentraciones adecuadas. La mejora solo funciona cuando las concentraciones de la ADN polimerasa 1 son mucho más bajas que los fragmentos de ADN que se van a ligar. Cuando las concentraciones de polimerasas de ADN Pol I son más altas, tiene un efecto adverso sobre la ligasa de ADN de E. coli.
T4
La ADN ligasa del bacteriófago T4 (un bacteriófago que infecta a la bacteria Escherichia coli). La ligasa T4 es la más utilizada en la investigación de laboratorio. Puede ligar extremos cohesivos o romos de ADN, oligonucleótidos, así como híbridos de ARN y ARN-ADN, pero no ácidos nucleicos monocatenarios. También puede ligar ADN de extremos romos con una eficacia mucho mayor que la ADN ligasa de E. coli. A diferencia de la ADN ligasa de E. coli, la ADN ligasa de T4 no puede utilizar NAD y tiene un requisito absoluto de ATP como cofactor. Se ha hecho algo de ingeniería para mejorar el in vitroactividad de T4 ADN ligasa; un enfoque exitoso, por ejemplo, probó la ligasa de ADN T4 fusionada con varias proteínas de unión de ADN alternativas y descubrió que las construcciones con p50 o NF-kB como socios de fusión eran más de un 160 % más activas en ligaciones de extremo romo con fines de clonación que el tipo salvaje T4 ADN ligasa. Una reacción típica para insertar un fragmento en un vector de plásmido usaría aproximadamente 0,01 (extremos cohesivos) a 1 (extremos romos) unidades de ligasa. La temperatura de incubación óptima para la ADN ligasa T4 es de 16 °C.
Los mutantes de ligasa del bacteriófago T4 tienen una mayor sensibilidad tanto a la radiación UV como al metanosulfonato de metilo del agente alquilante, lo que indica que la ligasa de ADN se emplea en la reparación de los daños en el ADN causados por estos agentes.
Mamífero
En los mamíferos, hay cuatro tipos específicos de ligasa.
- ADN ligasa I: liga el ADN naciente de la hebra rezagada después de que la Ribonucleasa H haya eliminado el cebador de ARN de los fragmentos de Okazaki.
- ADN ligasa III: forma complejos con la proteína de reparación de ADN XRCC1 para ayudar a sellar el ADN durante el proceso de reparación por escisión de nucleótidos y fragmentos recombinantes. De todas las ligasas de ADN de mamíferos conocidas, solo se ha encontrado que la Lig III está presente en las mitocondrias.
- ADN ligasa IV: complejos con XRCC4. Cataliza el paso final en el extremo no homólogo que une la ruta de reparación de roturas de doble cadena del ADN. También se requiere para la recombinación V(D)J, el proceso que genera diversidad en inmunoglobulinas y loci de receptores de células T durante el desarrollo del sistema inmunitario.
- ADN ligasa II: un artefacto de purificación resultante de la degradación proteolítica de la ADN ligasa III. Inicialmente, se ha reconocido como otra ADN ligasa y es la razón de la nomenclatura inusual de las ADN ligasas.
La ligasa de ADN de eucariotas y algunos microbios utiliza trifosfato de adenosina (ATP) en lugar de NAD.
Termoestable
Derivada de una bacteria termófila, la enzima es estable y activa a temperaturas mucho más altas que las ligasas de ADN convencionales. Su vida media es de 48 horas a 65 °C y superior a 1 hora a 95 °C. Se ha demostrado que Ampligase DNA Ligase es activa durante al menos 500 ciclos térmicos (94 °C/80 °C) o 16 horas de ciclo. Esta termoestabilidad excepcional permite una rigurosidad de hibridación y una especificidad de ligadura extremadamente altas.
Medición de la actividad
Hay al menos tres unidades diferentes que se utilizan para medir la actividad de la ADN ligasa:
- Unidad de Weiss: la cantidad de ligasa que cataliza el intercambio de 1 nmol de P de pirofosfato inorgánico a ATP en 20 minutos a 37 C. Esta es la más utilizada.
- Unidad Modrich-Lehman: rara vez se usa, y una unidad se define como la cantidad de enzima necesaria para convertir 100 nmoles de d(AT) n en una forma resistente a exonucleasa-III en 30 minutos en condiciones estándar.
- Muchos proveedores comerciales de ligasas utilizan una unidad arbitraria basada en la capacidad de la ligasa para ligar extremos cohesivos. Estas unidades suelen ser más subjetivas que cuantitativas y carecen de precisión.
Aplicaciones de investigación
Las ligasas de ADN se han convertido en herramientas indispensables en la investigación moderna de biología molecular para generar secuencias de ADN recombinante. Por ejemplo, las ligasas de ADN se utilizan con enzimas de restricción para insertar fragmentos de ADN, a menudo genes, en plásmidos.
Controlar la temperatura óptima es un aspecto vital para realizar experimentos de recombinación eficientes que impliquen la ligadura de fragmentos con extremos cohesivos. La mayoría de los experimentos utilizan ADN ligasa T4 (aislada del bacteriófago T4), que es más activa a 37 °C. Sin embargo, para una eficiencia de ligadura óptima con fragmentos con extremos cohesivos ("extremos cohesivos"), la temperatura óptima de la enzima debe equilibrarse con la temperatura de fusión T m de los extremos cohesivos que se ligan.el emparejamiento homólogo de los extremos cohesivos no será estable porque la alta temperatura interrumpe los enlaces de hidrógeno. Una reacción de ligadura es más eficaz cuando los extremos cohesivos ya están recocidos de manera estable y, por lo tanto, la ruptura de los extremos recocidos daría como resultado una baja eficiencia de ligadura. Cuanto más corto sea el voladizo, menor será la T m.
Dado que los fragmentos de ADN de extremos romos no tienen extremos cohesivos para hibridar, la temperatura de fusión no es un factor a considerar dentro del rango de temperatura normal de la reacción de ligadura. El factor limitante en la ligadura de extremos romos no es la actividad de la ligasa, sino el número de alineaciones entre los extremos de los fragmentos de ADN que se producen. Por lo tanto, la temperatura de ligadura más eficiente para el ADN de extremos romos sería la temperatura a la que puede ocurrir el mayor número de alineaciones. La mayoría de las ligaduras de extremos romos se realizan a 14-25 °C durante la noche. La ausencia de extremos recocidos de manera estable también significa que la eficiencia de la ligadura se reduce, lo que requiere el uso de una mayor concentración de ligasa.
Se puede ver un uso novedoso de la ADN ligasa en el campo de la nanoquímica, específicamente en el origami de ADN. Los principios de autoensamblaje basados en el ADN han resultado útiles para organizar objetos a nanoescala, como biomoléculas, nanomáquinas, componentes nanoelectrónicos y fotónicos. El ensamblaje de tal nanoestructura requiere la creación de una intrincada malla de moléculas de ADN. Aunque el autoensamblaje del ADN es posible sin ninguna ayuda externa utilizando diferentes sustratos, como la provisión de una superficie catatónica de papel de aluminio, la ligasa de ADN puede proporcionar la asistencia enzimática que se requiere para hacer una estructura de red de ADN a partir de fragmentos de ADN.
Historia
La primera ligasa de ADN fue purificada y caracterizada en 1967 por los laboratorios Gellert, Lehman, Richardson y Hurwitz. Primero fue purificado y caracterizado por Weiss y Richardson utilizando un proceso de fraccionamiento cromatográfico de seis pasos que comienza con la eliminación de los restos celulares y la adición de estreptomicina, seguido de varios lavados de columna de dietilaminoetil (DEAE)-celulosa y un fraccionamiento final de fosfocelulosa. El extracto final contenía el 10 % de la actividad registrada inicialmente en los medios de E. coli; a lo largo del proceso se descubrió que ATP y Mg++ eran necesarios para optimizar la reacción. Las ligasas de ADN comunes disponibles comercialmente se descubrieron originalmente en el bacteriófago T4, E. coli y otras bacterias.
Trastornos
Las deficiencias genéticas en las ADN ligasas humanas se han asociado con síndromes clínicos marcados por inmunodeficiencia, sensibilidad a la radiación y anomalías del desarrollo. El síndrome LIG4 (síndrome de la ligasa IV) es una enfermedad rara asociada con mutaciones en la ADN ligasa 4 e interfiere con los mecanismos de reparación de roturas del ADN bicatenario. El síndrome de ligasa IV causa inmunodeficiencia en individuos y se asocia comúnmente con microcefalia e hipoplasia medular. Una lista de enfermedades prevalentes causadas por la falta o el mal funcionamiento de la ADN ligasa es la siguiente.
Xeroderma pigmentoso
El xeroderma pigmentoso, comúnmente conocido como XP, es una afección hereditaria caracterizada por una sensibilidad extrema a los rayos ultravioleta (UV) de la luz solar. Esta condición afecta principalmente los ojos y las áreas de la piel expuestas al sol. Algunas personas afectadas también tienen problemas relacionados con el sistema nervioso.
Ataxia-telangiectasia
Las mutaciones en el gen ATM causan ataxia-telangiectasia. El gen ATM proporciona instrucciones para producir una proteína que ayuda a controlar la división celular y está involucrada en la reparación del ADN. Esta proteína juega un papel importante en el desarrollo y la actividad normal de varios sistemas del cuerpo, incluidos el sistema nervioso y el sistema inmunológico. La proteína ATM ayuda a las células a reconocer las hebras de ADN dañadas o rotas y coordina la reparación del ADN mediante la activación de enzimas que reparan las hebras rotas. La reparación eficiente de las hebras de ADN dañadas ayuda a mantener la estabilidad de la información genética de la célula. Los niños afectados suelen desarrollar dificultad para caminar, problemas con el equilibrio y la coordinación de las manos, movimientos espasmódicos involuntarios (corea), espasmos musculares (mioclono) y alteraciones en la función nerviosa (neuropatía). Los problemas de movimiento generalmente hacen que las personas requieran la asistencia de una silla de ruedas en la adolescencia. Las personas con este trastorno también tienen dificultad para hablar y dificultad para mover los ojos para mirar de lado a lado (apraxia oculomotora).
Anemia de Fanconi
La anemia de Fanconi (AF, por sus siglas en inglés) es un trastorno sanguíneo hereditario poco frecuente que provoca insuficiencia de la médula ósea. La AF evita que la médula ósea produzca suficientes células sanguíneas nuevas para que el cuerpo funcione normalmente. La FA también puede hacer que la médula ósea produzca muchas células sanguíneas defectuosas. Esto puede conducir a problemas de salud graves, como la leucemia.
Síndrome de Bloom
El síndrome de Bloom da como resultado una piel que es sensible a la exposición al sol y, por lo general, el desarrollo de un parche de piel enrojecida en forma de mariposa en la nariz y las mejillas. Una erupción cutánea también puede aparecer en otras áreas que normalmente están expuestas al sol, como el dorso de las manos y los antebrazos. Pequeños grupos de vasos sanguíneos agrandados (telangiectasias) a menudo aparecen en la erupción; las telangiectasias también pueden ocurrir en los ojos. Otras características de la piel incluyen parches de piel que son más claros o más oscuros que las áreas circundantes (hipopigmentación o hiperpigmentación, respectivamente). Estos parches aparecen en áreas de la piel que no están expuestas al sol y su desarrollo no está relacionado con las erupciones.
Como blanco de un fármaco
En estudios recientes, se usó ADN ligasa I humana en el diseño de fármacos asistido por computadora para identificar inhibidores de ADN ligasa como posibles agentes terapéuticos para tratar el cáncer. Dado que el crecimiento celular excesivo es un sello distintivo del desarrollo del cáncer, la quimioterapia dirigida que interrumpe el funcionamiento de la ligasa de ADN puede impedir las formas de cáncer adyuvantes. Además, se ha demostrado que las ADN ligasas se pueden dividir ampliamente en dos categorías, a saber, dependientes de ATP y NAD. Investigaciones anteriores han demostrado que, aunque se han descubierto ligasas de ADN dependientes de NAD en nichos celulares o virales esporádicos fuera del dominio bacteriano de la vida, no hay ningún caso en el que una NADLa ligasa dependiente de - está presente en un organismo eucariótico. La presencia únicamente en organismos no eucarióticos, la especificidad de sustrato única y la estructura de dominio distintiva de las ligasas de ADN humano dependientes de NAD+ en comparación con las dependientes de ATP hacen que las ligasas dependientes de NAD sean objetivos ideales para el desarrollo de nuevos fármacos antibacterianos.
Contenido relacionado
Ciclosis
Tilacoide
Endosoma