
11-desoxicorticosterona
La 11-desoxicorticosterona (DOC), o simplemente desoxicorticosterona, también conocida como 21-hidroxiprogesterona, así como desoxicorticosterona (DCI), desoxicorticosterona y cortexona, es una hormona esteroide producida por la glándula suprarrenal que posee actividad mineralocorticoide y actúa como precursor de la aldosterona. Es un mineralocorticoide activo (que retiene Na+). Como sus nombres lo indican, la 11-desoxicorticosterona puede entenderse como la variante 21-hidroxi de la progesterona o como la variante 11-desoxi de la corticosterona.
DOCA es la abreviatura del éster acetato de 11-desoxicorticosterona.
Actividad biológica
El DOC es un mineralocorticoide potente pero prácticamente no tiene actividad glucocorticoide. Sin embargo, la 11β-hidroxilación del DOC produce corticosterona y confiere actividad glucocorticoide, junto con una actividad mineralocorticoide diez veces menor. Además de su actividad mineralocorticoide, se ha descubierto que el DOC posee entre un tercio y un décimo de la potencia de la progesterona como progestágeno cuando se administra sistemáticamente a conejos. Sin embargo, no tiene dicha actividad cuando se aplica directamente a la mucosa uterina de ratones. La discrepancia puede estar relacionada con el hecho de que el DOC puede convertirse en progesterona in vivo.
Función biológica
La DOC es una molécula precursora de la producción de aldosterona. La vía principal de producción de aldosterona se encuentra en la zona glomerular de la glándula suprarrenal. No es una hormona secretora importante. Se produce a partir de la progesterona por acción de la 21β-hidroxilasa y se convierte en corticosterona por acción de la 11β-hidroxilasa. La corticosterona se convierte luego en aldosterona por acción de la aldosterona sintasa.
La mayor parte del DOC es secretado por la zona fasciculada de la corteza suprarrenal, que también secreta cortisol, y una pequeña cantidad por la zona glomerulosa, que secreta aldosterona. El DOC estimula los túbulos colectores (los túbulos que se ramifican para alimentar la vejiga) para que sigan excretando potasio de forma muy similar a como lo hace la aldosterona, pero no como la aldosterona en el extremo de los túbulos en forma de bucle (distal). Al mismo tiempo, no es tan riguroso en la retención de sodio como la aldosterona, más de 20 veces menos. El DOC representa solo el 1% de la retención de sodio normal. Además de su inherente falta de vigor, existe un mecanismo de escape controlado por una hormona no esteroide desconocida que anula el poder de conservación de sodio del DOC después de unos días, al igual que la aldosterona. Esta hormona puede ser la hormona peptídica calicreína, que es aumentada por el DOC y suprimida por la aldosterona. Si el sodio se eleva mucho, el DOC también aumenta el flujo de orina. El DOC tiene aproximadamente 1/20 del poder de retención de sodio de la aldosterona, y se dice que es tan poco como un uno por ciento de la aldosterona con una ingesta alta de agua. Dado que el DOC tiene aproximadamente 1/5 del poder de excreción de potasio de la aldosterona, probablemente debe contar con la ayuda de la aldosterona si el contenido de potasio sérico se vuelve demasiado alto. Las inyecciones de DOC no causan mucha excreción adicional de potasio cuando la ingesta de sodio es baja. Esto probablemente se debe a que la aldosterona ya está estimulando la salida de potasio. Cuando el sodio es bajo, el DOC probablemente no tendría que estar presente, pero cuando el sodio aumenta, la aldosterona disminuye considerablemente y el DOC probablemente tiende a tomar el control.
El DOC tiene una retroalimentación similar con respecto al potasio que la aldosterona. Un aumento del potasio sérico provoca un aumento de la secreción de DOC. Sin embargo, el sodio tiene poco efecto, y el efecto que tiene es directo. La angiotensina (la hormona de la presión arterial) tiene poco efecto sobre el DOC, pero el DOC provoca una rápida caída de la renina y, por lo tanto, de la angiotensina I, el precursor de la angiotensina II. Por lo tanto, el DOC debe estar inhibiendo indirectamente a la aldosterona, ya que la aldosterona depende de la angiotensina II. El sodio, y por lo tanto el volumen sanguíneo, es difícil de regular internamente. Es decir, cuando una gran dosis de sodio amenaza al cuerpo con presión arterial alta, no se puede resolver transfiriendo sodio al espacio intracelular (dentro de la célula). Los glóbulos rojos habrían sido posibles, pero eso no cambiaría el volumen sanguíneo. El potasio, por otro lado, puede ser trasladado al gran espacio intracelular, y aparentemente es mediante el DOC en conejos. Por lo tanto, un problema de potasio elevado en sangre puede resolverse en cierta medida sin tener que deshacerse de una cantidad excesiva de este mineral, que a veces es peligrosamente escaso y que no se puede bombear de forma activa independientemente del sodio. Es imperativo mantener un nivel adecuado de potasio total, ya que una deficiencia hace que el corazón pierda fuerza. El movimiento de potasio hacia las células intensificaría en cierta medida el problema del sodio, ya que cuando el potasio ingresa en la célula, sale una cantidad algo menor de sodio. Por lo tanto, es deseable resolver el problema de la presión arterial en la medida de lo posible mediante la disminución de la renina mencionada anteriormente, evitando así la pérdida de sodio, que normalmente era muy escaso en las sabanas africanas donde probablemente evolucionaron los antepasados humanos.
La semejanza del patrón de las fuerzas electromotrices producidas por la DOC en los túbulos renales con la ingesta normal de potasio, y la total disimilitud de su forma con respecto a la producida por túbulos deficientes en potasio, tenderían a apoyar la opinión anterior. Los atributos anteriores son consistentes con una hormona de la que se depende para descargar tanto el exceso de sodio como de potasio. La acción de la DOC para aumentar la calicreína, la hormona peptídica que se cree que es la "hormona de escape" del sodio, y la acción de la aldosterona para suprimirla, también respaldan el concepto anterior.
La ACTH tiene más efecto sobre la DOC que sobre la aldosterona. Esto puede ser para dar al sistema inmunológico el control sobre la regulación de electrolitos durante la diarrea, ya que durante la deshidratación, la aldosterona prácticamente desaparece aunque la renina y la angiotensina aumentan. Debido a que la aldosterona desaparece, los suplementos de potasio son muy peligrosos durante la deshidratación y no deben intentarse hasta al menos una hora después de la rehidratación para que las hormonas puedan llegar al núcleo.
El objetivo principal del DOC es regular los electrolitos. Sin embargo, tiene otros efectos, como eliminar el potasio de los leucocitos y los músculos, deprimir la formación de glucógeno y estimular la enzima lisiloxidasa que contiene cobre y el tejido conectivo, atributos que el cuerpo puede utilizar para ayudar a sobrevivir durante las enfermedades intestinales que provocan pérdida de potasio. La mayor eficiencia del DOC para permitir la excreción de sodio (o tal vez debería expresarse como ineficiencia en la retención) debe deberse en parte a cambios morfológicos en las células renales, ya que el escape de la retención de sodio del DOC tarda varios días en materializarse y, cuando lo hace, estas células son mucho más eficientes para descargar sodio si luego se agrega sodio que las células acostumbradas a una ingesta baja previa. Por lo tanto, paradójicamente, una ingesta baja de sal debería proteger contra la pérdida de sodio en la transpiración.
La progesterona previene parte de la pérdida de potasio causada por la DOC.
Imágenes adicionales
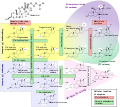 Steroidogenesis
Steroidogenesis Corticosterona
Corticosterona


Véase también
- 11-Deoxycortisol
- 17α-Hydroxyprogesterone
- Corticosterona
- Hiperplasia suprarrenal congénita
Referencias
- ^ Buckingham, MacDonald " Heilbron 1995.
- ^ Swiss Pharmaceutical Society 2011.
- ^ Costanzo 2014.
- ^ Harper Illustrated Biochemistry 30th Edition
- ^ Wang D, Luo Y, Myakala K, Orlicky DJ, Dobrinskikh E, Wang X, Levi M (agosto de 2017). "La serelaxin mejora la función cardíaca y renal en ratas hipertensivas con sal DOCA". Scientific Reports. 7 (1): 9793. Bibcode:2017NatSR...7.9793W. doi:10.1038/s41598-017-09470-0. PMC 5574886. PMID 28851937.
- ^ Lan NC, Graham B, Bartter FC, Baxter JD (febrero de 1982). "Binding of esteroides to mineralocorticoid receives: implications for in vivo occupancy by glucocorticoids". J Clin Endocrinol Metab. 54 (2): 332–42. doi:10.1210/jcem-54-2-332. PMID 6274900.
- ^ Funder JW (enero de 2005). "Receptores Mineralocorticoides: distribución y activación". Heart Fail Rev. 10 (1): 15–22. doi:10.1007/s10741-005-2344-2. PMID 15947887. S2CID 1423344.
- ^ a b Goodman HM (28 de julio de 2010). Endocrinología médica básica. Academic Press. p. 64. ISBN 978-0-08-092055-9.
- ^ a b c Las hormonas adrenocorticales: su origen · Química, Fisiología y Farmacología. Springer Science & Business Media. 27 noviembre 2013. p. 610. ISBN 978-3-642-88385-9.
- ^ Lieberman, Marks & Peet 2012.
- ^ a b c O'Neil " Helman 1977.
- ^ Peterson " Wright 1977.
- ^ Ellinghaus 1971.
- ^ Ruch " Fulton 1960.
- ^ Pearce et al. 1969.
- ^ Schacht, Lowenstein & Baldwin 1971.
- ^ Majima et al. 1999.
- ^ a b Bönner et al. 1981.
- ^ a b c Oddie, Coghlan " Scoggins 1972.
- ^ Desaulles 1958.
- ^ Bauer & Gauntner 1979.
- ^ a b Brown, Strott & Liddle 1972.
- ^ Schambelan & Biglieri 1972.
- ^ a b Grekin, Terris " Bohr 1980.
- ^ Abrecht 1972.
- ^ Rubini & Chojnacki 1972.
- ^ Merrill, Skelton & Cowley 1986.
- ^ Wilson 1957.
- ^ Tobian & Binion 1954.
- ^ Bartlett & MacKay 1949.
- ^ Pospísilová " Pospísil 1970.
- ^ Wambach " Higgins 1979.
Fuentes
- Abrecht PH (septiembre de 1972). "Efectos cardiovasculares de deficiencia crónica de potasio en el perro". El American Journal of Physiology. 223 (3): 555-60. doi:10.1152/ajplegacy.1972.223.3.555. PMID 4403425.
- Bartlett GR, MacKay EM (julio de 1949). "Estimulación de insulina de la formación de glucógeno en el músculo abdominal de rata". Actas de la Sociedad de Biología Experimental y Medicina. 71 (3): 493-5. doi:10.3181/00379727-71-17234. PMID 18136520. S2CID 31824669.
- Bauer JH, Gauntner WC (marzo de 1979). "Efecto del cloruro de potasio en la actividad de renina de plasma y aldosterona de plasma durante la restricción de sodio en el hombre normal". Kidney International. 15 (3): 286–93. doi:10.1038/ki.1979.37. PMID 513492.
- Bönner G, Autenrieth R, Marin-Grez M, Rascher W, Gross F (1981). "Efectos de carga de sodio, acetato de desoxycorticosterona y corticosterona en excreción urinaria de kallikrein". Investigación hormonal. 14 (2): 87–94. doi:10.1159/000179365 (inactivo 22 de junio de 2024). PMID 6912177.
{{cite journal}}: CS1 maint: DOI inactive as of June 2024 (link) - Brown RD, Strott CA, Liddle GW (junio de 1972). "Site of stimulation of aldosterone biosynthesis by angiotensin and potassium". The Journal of Clinical Investigation. 51 (6): 1413-8. doi:10.1172/JCI106937. PMC 292278. PMID 4336939.
- Buckingham J, MacDonald F, Heilbron I, eds. (1995). Diccionario de compuestos orgánicosVol. 1 (6a edición). CRC Prensa. ISBN 978-0412540905.
- Costanzo LS (2014). Fisiología (International ed.). Lippincott Williams & Wilkins. ISBN 978-1469832005.
{{cite book}}:|work=ignorado (ayuda) - Desaulles P (1958). Muller AF, O'Connor CM (eds.). Comparación de los efectos de la aldosterona, la cortexona y el cortisol en ratas adrenalectomizadas bajo diversas cargas de sal. An International Symposium on Aldosterone, Geneva 1957. Boston: Little Brown Co. pp. 29–38. OCLC 14594852.
- Ellinghaus K (1971). "El equilibrio entre sodio y potasio durante la administración de desoxycorticosterona en perros con diferentes ingestas dietéticas de sodio". Pflügers Archiv. 322 (4): 347-54. doi:10.1007/BF00587752. PMID 5100181. S2CID 1968289.
- Grekin RJ, Terris JM, Bohr DF (1980). "Efectos electrolitos y hormonales de acetato de deoxycorticosterona en cerdos jóvenes". Hipertensión. 2 (3): 326-32. doi:10.1161/01.HYP.2.326. PMID 6993359.
- Lieberman MA, Marks A, Peet A (2012). Médico básico de Marks Bioquímica: Enfoque clínico. Matthew Chansky (Illustrator) (4th ed.). Lippincott Williams y Wilkins. ISBN 978-1608315727.
- Majima M, Hayashi I, Fujita T, Ito H, Nakajima S, Katori M (octubre de 1999). "La facilitación del sistema de kallikrein-kinin renal evita el desarrollo de la hipertensión por la inhibición de la retención de sodio". Inmunopharmacology. 44 (1–2): 145–52. doi:10.1016/S0162-3109(99)00086-7. PMID 10604538.
- Merrill DC, Skelton MM, Cowley AW (junio de 1986). "El control humano del agua y la excreción electrolítica durante la restricción del agua". Kidney International. 29 (6): 1152 a 61. doi:10.1038/ki.1986.121. PMID 3528610.
- Oddie CJ, Coghlan JP, Scoggins BA (junio de 1972). "Los niveles de plasma desoxycorticosterona en el hombre con medición simultánea de aldosterona, corticosterona, cortisol y 11-deoxycortisol". The Journal of Clinical Endocrinology and Metabolism. 34 (6): 1039–54. doi:10.1210/jcem-34-6-1039. PMID 4336456.
- O'Neil RG, Helman SI (diciembre de 1977). "Características de transporte de tubulas recogiendo renales: influencias de DOCA y dieta". El American Journal of Physiology. 233 (6): F544-58. doi:10.1152/ajprenal.1977.233.6.F544. PMID 596453.
- Pearce JW, Sonnenberg H, Veress AT, Ackermann U (abril de 1969). "Evidencia para un factor humoral que modifica la respuesta renal a la expansión del volumen sanguíneo en la rata". Canadian Journal of Physiology and Pharmacology. 47 (4): 377–86. doi:10.1139/y69-066. PMID 5772633.
- Peterson LN, Wright FS (septiembre 1977). "Efecto de la ingesta de sodio en la excreción renal de potasio". El American Journal of Physiology. 233 (3): F225-34. doi:10.1152/ajprenal.1977.233.3.F225. PMID 910918.
- Pospísilová J, Pospísil M (1970). "Influencia de mineralocorticoides en síntesis de colágeno en granuloma subcutáneo en ratones adrenalectomizados y no adrenalectomizados". Fisiología Bohemoslovaca. 19 (6): 539–43. PMID 4251388.
- Rubini ME, Chojnacki RE (enero de 1972). "Principios de terapia parenteral". El American Journal of Clinical Nutrition. 25 (1): 96–113. doi:10.1093/ajcn/25.1.96. PMID 5007603.
- Ruch T, Fulton JF, eds. (1960). Fisiología médica y biofísica. W. B. Saunders and Co.
- Schacht RG, Lowenstein J, Baldwin DS (octubre de 1971). "Mecanismo real para el escape de DOCA en el hombre". Boletín de la Academia de Medicina de Nueva York. 47 10): 1233. PMID 5286466.
- Schambelan M, Biglieri EG (abril de 1972). "Producción y regulación de la deoxicorticosterona en el hombre". The Journal of Clinical Endocrinology and Metabolism. 34 (4): 695–703. doi:10.1210/jcem-34-4-695. PMID 4335105.
- Swiss Pharmaceutical Society (2011). Índice Nominum: Directorio Internacional de Drogas (20a edición). Wissenschaftliche Verlag. ISBN 978-3804750531.
- Tobian L, Binion J (octubre de 1954). "E electrolitos de pared de arte en hipertensión renal y DCA". The Journal of Clinical Investigation. 33 (10): 1407-14. doi:10.1172/JCI103018. PMC 1072561. PMID 13201647.
- Wambach G, Higgins JR (marzo de 1979). "Efecto de progesterona en concentración de electrolitos de suero y tejido en ratas tratadas por doca". Investigación hormonal y metabólica. 11 (3): 258-9. doi:10.1055/s-0028-1095777. PMID 447212. S2CID 36911377.
- Wilson DL (Julio 1957). "Efectos directos de los esteroides corticales suprarrenales en el contenido de electrolitos de leucocitos de conejo". El American Journal of Physiology. 190 (1): 104–8. doi:10.1152/ajplegacy.1957.190.1.104. PMID 13458419.