
Evolução convergente


Evolução convergente é a evolução independente de características semelhantes em espécies de diferentes períodos ou épocas no tempo. A evolução convergente cria estruturas análogas que têm forma ou função semelhante, mas não estavam presentes no último ancestral comum desses grupos. O termo cladístico para o mesmo fenômeno é a homoplasia. A evolução recorrente do vôo é um exemplo clássico, pois insetos voadores, pássaros, pterossauros e morcegos evoluíram independentemente a capacidade útil do voo. Recursos funcionalmente semelhantes que surgiram através da evolução convergente são análogos , enquanto as estruturas ou características de homólogas têm uma origem comum, mas podem ter funções diferentes. As asas de pássaros, morcegos e pterossauros são estruturas análogas, mas seus membros anteriores são homólogos, compartilhando um estado ancestral, apesar de cumprir funções diferentes.
O oposto da convergência é a evolução divergente, onde espécies relacionadas evoluem diferentes características. A evolução convergente é semelhante à evolução paralela, que ocorre quando duas espécies independentes evoluem na mesma direção e, portanto, adquirem independentemente características semelhantes; Por exemplo, os sapos deslizantes evoluíram em paralelo de vários tipos de sapo de árvore.
Muitos casos de evolução convergente são conhecidos em plantas, incluindo o desenvolvimento repetido da fotossíntese C4, a dispersão de sementes por frutas carnudas adaptadas para serem comidas por animais e carnivoria.
Visão geral

Na morfologia, traços análogos surgem quando espécies diferentes vivem de maneiras semelhantes e/ou um ambiente semelhante, e assim enfrentam os mesmos fatores ambientais. Ao ocupar nichos ecológicos semelhantes (ou seja, um modo de vida distinto) problemas semelhantes podem levar a soluções semelhantes. O anatomista britânico Richard Owen foi o primeiro a identificar a diferença fundamental entre analogias e homologias.
Em bioquímica, restrições físicas e químicas nos mecanismos causaram alguns acordos ativos do local, como a tríade catalítica, evoluirem independentemente em superfamílias enzimáticas separadas.
Em seu livro de 1989 vida maravilhosa , Stephen Jay Gould argumentou que, se alguém pudesse rebobinar a fita da vida [e] as mesmas condições foram encontradas novamente, a evolução poderia levar uma muito diferente Curso. " Simon Conway Morris contesta essa conclusão, argumentando que a convergência é uma força dominante na evolução e, dado que as mesmas restrições ambientais e físicas estão em ação, a vida inevitavelmente evoluirá em direção a um " otimum " Plano corporal e, em algum momento, a evolução deve tropeçar na inteligência, uma característica atualmente identificada com pelo menos primatas, corvidas e cetáceos.
Distinções
Cladistics
Na cladística, uma homoplasia é uma característica compartilhada por dois ou mais táxons por qualquer motivo que não seja uma ascendência comum. Os táxons que compartilham ancestralidade fazem parte do mesmo clado; A cladística procura organizá -los de acordo com seu grau de parentesco para descrever sua filogenia. As características homoplásticas causadas pela convergência são, portanto, do ponto de vista da cladística, fatores de confusão que podem levar a uma análise incorreta.
Atavismo
Em alguns casos, é difícil dizer se uma característica foi perdida e depois re evoluiu de forma convergente, ou se um gene simplesmente foi desligado e depois reativado posteriormente. Uma característica tão ressurgida é chamada de atavismo. Do ponto de vista matemático, um gene não utilizado (seletivamente neutro) tem uma probabilidade de retenção constantemente de retenção de funcionalidade potencial ao longo do tempo. A escala de tempo desse processo varia muito em diferentes filogenias; Em mamíferos e pássaros, há uma probabilidade razoável de permanecer no genoma em um estado potencialmente funcional por cerca de 6 milhões de anos.
Evolução paralela versus convergente

Quando duas espécies são semelhantes em um caráter específico, a evolução é definida como paralela se os ancestrais também fossem semelhantes e convergentes se não fossem. Alguns cientistas argumentaram que há um continuum entre evolução paralela e convergente, enquanto outros sustentam que, apesar de alguma sobreposição, ainda existem distinções importantes entre os dois.
Quando as formas ancestrais não são especificadas ou desconhecidas, ou a gama de características consideradas não é claramente especificada, a distinção entre evolução paralela e convergente se torna mais subjetiva. Por exemplo, o exemplo impressionante de formas placentárias e marsupiais semelhantes é descrito por Richard Dawkins em o relojoeiro cego como um caso de evolução convergente, porque os mamíferos em cada continente tinham uma longa história evolutiva antes da extinção de os dinossauros sob os quais acumularem diferenças relevantes.
A nível molecular

Proteínas
Protease sites ativos
A enzimologia das proteases fornece alguns dos exemplos mais claros de evolução convergente. Esses exemplos refletem as restrições químicas intrínsecas nas enzimas, levando a evolução a convergir em soluções equivalentes de forma independente e repetidamente.
As proteases de serina e cisteína usam diferentes grupos funcionais de aminoácidos (álcool ou tiol) como nucleófilo. Para ativar esse nucleófilo, eles orientam um resíduo ácido e básico em uma tríade catalítica. As restrições químicas e físicas na catálise enzimática causaram arranjos idênticos à tríade para evoluir independentemente mais de 20 vezes em diferentes superfamílias enzimáticas. As proteases de treonina usam o aminoácido treonina como seu nucleófilo catalítico. Ao contrário da cisteína e da serina, a treonina é um álcool secundário (ou seja, possui um grupo metil). O grupo metil de treonina restringe bastante as possíveis orientações da tríade e do substrato, à medida que os metil se chocam com o backbone da enzima ou a base da histidina. Consequentemente, a maioria das proteases de treonina usa um treonina N-terminal para evitar tais confrontos estéricos. Várias superfamílias enzimáticas evolutivamente independentes com diferentes dobras de proteínas usam o resíduo N-terminal como nucleófilo. Essa semelhança do local ativo, mas a diferença de dobra de proteínas indica que o local ativo evoluiu de forma convergente nessas famílias.Cone snail e peixe insulina
Conus geographus produz uma forma distinta de insulina que é mais semelhante às sequências de proteínas de insulina de peixe do que à insulina de moluscos mais intimamente relacionados, sugerindo evolução convergente, embora com a possibilidade de transferência horizontal de genes.
A absorção de ferro ferro ferro ferro ferro através de transportadores de proteínas em plantas terrestres e clorofitos
homólogos distantes dos transportadores de íons metálicos zip em plantas terrestres e clorófitos convergiram em estrutura, provavelmente adotar Fe
Na+, K+-ATPase e resistência do inseto a esteróides cardiotônicos
Muitos exemplos de evolução convergente existem em insetos em termos de desenvolvimento de resistência em nível molecular a toxinas. Um exemplo bem caracterizado é a evolução da resistência a esteróides cardiotônicos (CTSs) via substituições de aminoácidos em posições bem definidas da subunidade α da Na+, K+-ATPase (Atpalpha). A variação no Atpalpha foi pesquisada em várias espécies adaptadas ao CTS, abrangendo seis ordens de insetos. Entre 21 espécies adaptadas ao CTS, 58 (76%) de 76 substituições de aminoácidos em locais implicados na resistência ao STC ocorrem em paralelo em pelo menos duas linhagens. 30 dessas substituições (40%) ocorrem em apenas dois locais na proteína (posições 111 e 122). As espécies adaptadas ao CTS também desenvolveram recorrente duplicações neofuncionalizadas de Atpalpha, com padrões de expressão específicos para tecidos convergentes.
Ácidos nucleicos
A convergência ocorre no nível do DNA e nas sequências de aminoácidos produzidas pela tradução de genes estruturais em proteínas. Estudos encontraram convergência em sequências de aminoácidos em morcegos ecolocantes e no golfinho; entre mamíferos marinhos; entre pandas gigantes e vermelhos; e entre o tilacino e os canídeos. A convergência também foi detectada em um tipo de DNA não codificante, elementos reguladores cis, como em suas taxas de evolução; Isso pode indicar seleção positiva ou seleção de purificação relaxada.
Na morfologia animal

Planos de corpo
natação animais, incluindo peixes como arenques, mamíferos marinhos, como golfinhos e ictiossauros (do mesozóico), todos convergiram na mesma forma simplificada. Uma forma semelhante e adaptações de natação estão presentes em moluscos, como Phylliroe . A forma de corpo fusiforme (um tubo diminuiu nas duas extremidades) adotada por muitos animais aquáticos é uma adaptação para permitir que viajassem em alta velocidade em um ambiente de alto arrasto. Formas do corpo semelhantes são encontradas nas vedações sem ouvidas e nas vedações de orelhas: elas ainda têm quatro pernas, mas elas são fortemente modificadas para nadar.
A fauna marsupial da Austrália e os mamíferos placentários do Velho Mundo têm várias formas surpreendentemente semelhantes, desenvolvidas em dois clados, isoladas uma da outra. O corpo, e especialmente a forma do crânio, do tilacino (tigre da Tasmânia ou lobo da Tasmânia) convergiu com os de Canidae como a raposa vermelha, vulpes vulpes .
- Convergência de mamíferos marsupiais e placentários
 esqueleto de raposa vermelha
esqueleto de raposa vermelha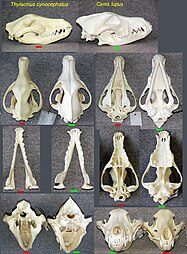 Caveiras de tilacina (esquerda), lobo de madeira (direita)
Caveiras de tilacina (esquerda), lobo de madeira (direita)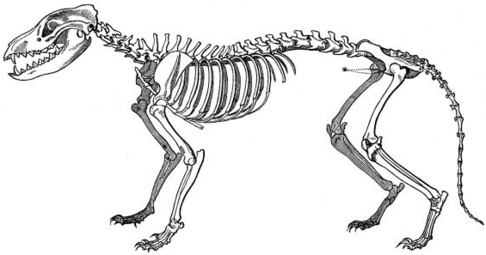 esqueleto de Thylacine
esqueleto de Thylacine


.png)
Ecologia
Como adaptação sensorial, o ecolocalização evoluiu separadamente em cetáceos (golfinhos e baleias) e morcegos, mas das mesmas mutações genéticas.
Peixes elétricos
Os gintotiformes da América do Sul e os Mormyridae da África evoluíram independentemente a eletrogerecepção passiva (cerca de 119 e 110 milhões de anos atrás, respectivamente). Cerca de 20 milhões de anos após a aquisição dessa habilidade, ambos os grupos desenvolveram a eletrogênese ativa, produzindo campos elétricos fracos para ajudá -los a detectar presas.
- Convergência de peixes fracamente elétricos
 Forma de onda de eletrolocalização de ginástica
Forma de onda de eletrolocalização de ginástica Um peixe elétrico de ginástica da América do Sul
Um peixe elétrico de ginástica da América do Sul Um peixe elétrico mormyrid da África
Um peixe elétrico mormyrid da África- Forma de onda de eletrolocalização Mormyrid


Olhos

Um dos exemplos mais conhecidos de evolução convergente é o olho da câmera de cefalópodes (como Squid e Octopus), vertebrados (incluindo mamíferos) e cnidaria (como água-viva). Seu último ancestral comum tinha, no máximo, um ponto fotorreceptivo simples, mas uma variedade de processos levou ao refinamento progressivo dos olhos da câmera - com uma diferença acentuada: o olho do cefalópode é "Wired " Na direção oposta, com os vasos sanguíneos e nervosos entrando na parte de trás da retina, em vez da frente como nos vertebrados. Como resultado, os cefalópodes não têm um ponto cego.
Voo

Hummingbird Hawk-Moths e Hummingbirds desenvolveram padrões semelhantes de voo e alimentação.
Partes da boca do inseto
Peças bucais de insetos mostram muitos exemplos de evolução convergente. As peças bucais de diferentes grupos de insetos consistem em um conjunto de órgãos homólogos, especializados na ingestão alimentar desse grupo de insetos. Evolução convergente de muitos grupos de insetos liderados de peças bucais originais de mordaça para diferentes tipos de funções derivadas. Isso inclui, por exemplo, a probóscide de insetos visitantes de flores, como abelhas e besouros de flores, ou as peças bucais que morrem de insetos sugadores de sangue, como pulgas e mosquitos.
Opposable polegares
Polegares opositados, permitindo que a compreensão dos objetos sejam mais frequentemente associados a primatas, como seres humanos, macacos, macacos e lêmures. Os polegares opositáveis também evoluíram em pandas gigantes, mas estes são completamente diferentes em estrutura, com seis dedos, incluindo o polegar, que se desenvolve a partir de um osso do pulso totalmente separadamente de outros dedos.
Primários
 Apesar do clareamento semelhante da cor da pele após sair da África, diferentes genes foram envolvidos nas linhagens européias (esquerda) e da Ásia Oriental (direita). Apesar do clareamento semelhante da cor da pele após sair da África, diferentes genes foram envolvidos nas linhagens européias (esquerda) e da Ásia Oriental (direita). | ||


_%EC%A0%9C%EC%9E%91%EA%B8%B0%EC%98%81%EC%83%81_%EC%97%84%EC%A0%95%ED%99%94_3m3s.jpg)
| Humanos | Lemurs | ||
|---|---|---|---|
| Apesar da semelhança da aparência, a base genética dos olhos azuis é diferente em humanos e lêmures. | |||

.jpg)
.jpg)

lêmures e humanos são primatos. Os primatas ancestrais tinham olhos castanhos, como a maioria dos primatas hoje. A base genética dos olhos azuis em humanos foi estudada em detalhes e muito se sabe sobre isso. Não é o caso de um locus de gene ser responsável, digamos, com a cor marrom dominante à cor dos olhos azuis. No entanto, um único locus é responsável por cerca de 80% da variação. Nos lêmures, as diferenças entre os olhos azuis e castanhos não são completamente conhecidos, mas o mesmo locus de gene não está envolvido.
Em plantas

O ciclo de vida anual
Enquanto a maioria das espécies de plantas é perene, cerca de 6% seguem um ciclo de vida anual, vivendo por apenas uma estação de crescimento. O ciclo de vida anual emergiu independentemente em mais de 120 famílias de angiospermas de plantas. A prevalência de espécies anuais aumenta em condições de verão a quente nas quatro famílias ricas em espécies de anuários (Asteraceae, Brassicaceae, Fabaceae e Poaceae), indicando que o ciclo de vida anual é adaptável.
Fixação de carbono
A fotossíntese C4, um dos três principais processos bioquímicos de fixação de carbono, surgiu independentemente de 40 vezes. Cerca de 7.600 espécies vegetais de angiospermas usam a fixação de carbono C , com muitos monocotes, incluindo 46% das gramíneas, como milho e cana -de -açúcar, e dicots, incluindo várias espécies no Chenopodiaceae e no Amaranthaceae.
Frutas
frutas com uma ampla variedade de origens estruturais convergiram para se tornarem comestíveis. Maçãs são pomes com cinco carpelos; Seus tecidos acessórios formam o núcleo da Apple, cercado por estruturas de fora da fruta botânica, o receptáculo ou hipontio. Outras frutas comestíveis incluem outros tecidos vegetais; A parte carnuda de um tomate é as paredes do pericarpo. Isso implica evolução convergente sob pressão seletiva, neste caso a competição por dispersão de sementes por animais através do consumo de frutas carnudas.
A dispersão de sementes por formigas (Myrmecochory) evoluiu independentemente mais de 100 vezes e está presente em mais de 11.000 espécies de plantas. É um dos exemplos mais dramáticos de evolução convergente na biologia.
Carnívoro

O carnivório evoluiu várias vezes independentemente em plantas em grupos amplamente separados. Em três espécies estudadas, cefalotus folicularis , nepenthes alata e Sarracenia purpurea , houve convergência no nível molecular. As plantas carnívoras secretam enzimas no fluido digestivo que produzem. Ao estudar fosfatase, glicosídeo hidrolase, glucanase, enzimas RNase e quitinase, bem como uma proteína relacionada à patogênese e uma proteína relacionada à taumatina, os autores encontraram muitas substituições de aminoácidos convergentes. Essas mudanças não foram nas enzimas ' Locais catalíticos, mas nas superfícies expostas das proteínas, onde podem interagir com outros componentes da célula ou do fluido digestivo. Os autores também descobriram que genes homólogos na planta não carnívoro Arabidopsis thaliana tendem a ter sua expressão aumentada quando a planta é estressada, levando os autores a sugerir que as proteínas responsivas ao estresse foram frequentemente cooptadas na evolução repetida do carnivório.
Métodos de inferência

Medidas baseadas em padrões
Métodos anteriores para medir a convergência incorporam proporções de distância fenotípica e filogenética, simulando a evolução com um modelo de movimento browniano de evolução de características ao longo de uma filogenia. Métodos mais recentes também quantificam a força da convergência. Uma desvantagem a ter em mente é que esses métodos podem confundir estase de longo prazo com convergência devido a semelhanças fenotípicas. A estase ocorre quando há pouca mudança evolutiva entre os táxons.
Medidas baseadas em distância avaliam o grau de similaridade entre as linhagens ao longo do tempo. As medidas baseadas em frequência avaliam o número de linhagens que evoluíram em um espaço de característica específico.
Medidas baseadas em processos
Métodos Para inferir modelos de seleção de ajuste de convergência baseados no processo para uma filogenia e dados de características contínuas para determinar se as mesmas forças seletivas agiram em linhagens. Isso usa o processo Ornstein -Uhlenbeck para testar diferentes cenários de seleção. Outros métodos dependem de uma especificação a priori de onde ocorreram mudanças na seleção.
Ver também
- Seleção incompleta de linhagem – Característica da análise filogenética: a presença de múltiplos alelos em populações ancestrais pode levar à impressão de que a evolução convergente ocorreu.
- Carcinização – Evolução de crustáceos em formas semelhantes a caranguejos
- Morfologia (biologia) – Estudo de formas externas e estruturas de organismos
- Evolução iterativa – A evolução repetida de um traço específico ou plano corporal da mesma linhagem ancestral em diferentes pontos no tempo.
- Elvis taxon – Misidentificação de posterior taxon superficialmente semelhante ao anterior extinto taxon
- Raça de volta – Uma forma de reprodução seletiva para recriar os traços de uma espécie extinta, mas o genoma difere da espécie original.
- Ortogênese (contraestável com evolução convergente; envolve teleologia)
- Contingência (biologia evolutiva) – efeito da história evolutiva sobre os resultados
Notas
- ^ No entanto, a biologia evolutiva do desenvolvimento identificou profunda homologia entre inseto e planos corporais mamíferos, para a surpresa de muitos biólogos.
- ^ No entanto, todos os organismos compartilham um ancestral comum mais ou menos recentemente, de modo que a questão de quão longe olhar no tempo evolutivo e como os ancestrais precisam ser semelhantes para se considerar a evolução paralela ter ocorrido não é inteiramente resolvida dentro da biologia evolutiva.
- ^ A existência prévia de estruturas adequadas foi chamada pré-adaptação ou exaptação.
Referências
- ^ Kirk, John Thomas Osmond (2007). Ciência e Certeza. Csiro Publishing. p. 79. ISBN 978-0-643-09391-1. Arquivado do original em 15 de fevereiro de 2017. Retrieved 23 de Janeiro 2017.
convergência evolutiva, que, citando.. Simon Conway Morris.. é a tendência de "recorrer da organização biológica para chegar à mesma "solução" a uma determinada "necessidade"... o " tigre tasmaniano" .. olhou e se comportava como um lobo e ocupou um nicho ecológico semelhante, mas na verdade era um marsupial não um mamífero placentário.
- ^ Reece, J.; Meyers, N.; Urry, L.; Cain, M.; Wasserman, S.; Minorsky, P.; Jackson, R.; Cooke, B. (5 de setembro de 2011). Cambell Biology, 9a Edição. Pearson. p. 586. ISBN 978-1-4425-3176-5.
- ↑ a b «Homologies and analogies» (em inglês). Universidade da Califórnia Berkeley. Arquivado do original em 19 de novembro de 2016. Retrieved 10 de Janeiro 2017.
- ^ Thunstad, Erik (2009). Darwins teori, evolusjon gjennom 400 anos (em norueguês). Oslo, Norway: Humanist forlag. p. 404. ISBN 978-82-92622-53-7.
- ↑ a b d Buller, A. R.; Townsend, C. A. (19 de fevereiro de 2013). «Intrinsic evolutionary restrings on protease structure, enzima acylation, and the identity of the catalytic triad» (em inglês). Actividades da Academia Nacional de Ciências dos Estados Unidos da América. 110 (8): E653–61. Bibcode:2013 PNAS.110E.653B. doi:10.1073/pnas.1221050110. PMC 35819. PMID 23382230.
- ^ Gould, S.J. (1989). Vida maravilhosa: o Burgess Shale e a natureza da história. W.W. Norton. pp. 282–285. ISBN 978-0-09-174271-3.
- ↑ a b c Conway Morris, Simon (2005). A solução da vida: seres humanos inevitáveis num universo solitário. Cambridge University Press. pp. 164, 167, 170 e 235. ISBN 978-0-521-60325-6. OCLC 156902715.
- ^ Chirat, R.; Moulton, D. E.; Goriely, A. (2013). «Base mecânica da morfogênese e evolução convergente das conchas marinhas espinhais». Procedimentos da Academia Nacional de Ciências. 110 (15): 6015–6020. Bibcode:2013PNAS..110.6015C. doi:10.1073/pnas.1220443110. PMC 362536. PMID 23530223.
- ^ Lomolino, M; Riddle, B; Whittaker, R; Brown, J (2010). Biogeografia, Quarta Edição. Sinauer Associates. p. 426. ISBN 978-0-87893-494-2.
- ^ West-Eberhard, Mary Jane (2003). Plasticidade e Evolução do Desenvolvimento. pp. 353–376. ISBN 978-0-19-512235-0.
- ^ Sanderson, Michael J.; Hufford, Larry (1996). Homoplasy: The Recurrence of Similarity in Evolution. Academic Press. pp. 330, e passim. ISBN 978-0-08-053411-4. Arquivado do original em 14 de fevereiro de 2017. Retrieved 21 de Janeiro 2017.
- ^ Collin, R.; Cipriani, R. (2003). «Dollo's law and the re-evolution of shell coiling» (em inglês). Processos da Royal Society B. 270 (1533): 2551–2555. doi:10.1098/rspb.2003.2517. PMC 1691546. PMID 14728776.
- ^ Arendt, J; Reznick, D (janeiro de 2008). "Convergência e paralelismo reconsiderados: o que aprendemos sobre a genética da adaptação?". Tendências em Ecologia e Evolução. 23 (1): 26–32. doi:10.1016/j.tree.2007.09.011. PMID 18022278.
- ^ Waters, Jonathan M.; McCulloch, Graham A. (2021). "Reinventar a roda? Reassessando os papéis do fluxo genético, classificação e convergência na evolução repetida". Ecologia molecular. 30 (17): 4162–4172. Bibcode:2021MolEc..30.4162W. doi:10.1111/mec.16018. ISSN 1365-294X. PMID 34133810. S2CID 235460165.
- ^ Pearce, T. (10 de novembro de 2011). «Convergence and Parallelism in Evolution: A Neo-Gouldian Account» (em inglês). O Jornal Britânico para a Filosofia da Ciência. 63 (2): 429–448.10.1093/bjps/axr046.
- ^ Zhang, J.; Kumar, S. (1997). «Detecção da evolução convergente e paralela no nível de sequência de aminoácidos». Mol. Evo. 14 (5): 527–36.10.1093/oxfordjournals.molbev.a025789. PMID 9159930.
- ^ Dawkins, Richard (1986). O Guardião Blind. W. W. Norton. pp. 100–106. ISBN 978-0-393-31570-7.
- ^ Dodson, G.; Wlodawer, A. (setembro de 1998). «Catalytic triads and their parents» (em inglês). Tendências em Ciências Bioquímicas. 23 (9): 347–52. doi:10.1016/S0968-0004(98)01254-7. PMID 9787641.
- ^ Ekici, O. D.; Paetzel, M.; Dalbey, R. E. (dezembro de 2008). «Unconventional serine proteases: changes on the catalytic Ser/His/Asp triad Configuration» (em inglês). Ciência da proteína. 17. (12): 2023–37. doi:10.1110/ps.035436.108. PMC 2590910. PMID 18824507.
- ^ Safavi-Hemami, Helena; Gajewiak, Joanna; Karanth, Santhosh; Robinson, Samuel D.; Ueberheide, Beatrix; Douglass, Adam D.; Schlegel, Amnon; Imperial, Julita S.; Watkins, Maren; Bandyopadhyay, Pradip K.; Yandell, Mark; Li, Qing; Norton Larga. «Specialized insulin is used for chemical war by fish-hunting cone snails» (em inglês). Procedimentos da Academia Nacional de Ciências. 112 (6): 1743–1748. Bibcode:2015PNAS..112.1743S. doi:10.1073/pnas.1423857112. PMC 4330763. PMID 25605914.
- ^ Martin, J. P.; Fridovich, I (1981). «Evidence for a natural gene-transfer from the ponyfish to its bioluminescent bacterial symbiont Photobacter leiognathi — the close relação between bacteriocuprein and the Copper-zinc superoxide-dismutase of teleost fishes» (em inglês). J. Biol. Chem. 256. (12): 6080–6089.10.1016/S0021-9258(19)69131-3. PMID 6787049.
- ^ Rodrigues, Wenderson Felipe Costa; Lisboa, Ayrton Breno P.; Lima, Joni Esrom; Ricachenevsky, Felipe Klein; Del-Bem, Luiz-Eduardo (10 de janeiro de 2023). «Ferrous iron uptake via IRT1 / ZIP evoluiu pelo menos duas vezes em plantas verdes». Novo médico. 237 (6): 1951-1961. - Sim.10.1111/nph.18661. PMID 36626937.
- ↑ a b Zhen, Ying; Aardema, Matthew L.; Medina, Edgar M.; Schumer, Molly; Andolfatto, Peter (28 de setembro de 2012). «Parallel Molecular Evolution in an Herbivore Community» (em inglês). Ciência. 337 (6102): 1634–1637. Código Bíblico:2012Sci...337.1634Z. doi:10.1126/science.1226630. ISSN 0036-8075. PMC 3770729. PMID 23019645.
- ^ Dobler, S., Dalla, S., Wagschal, V., & Agrawal, A. A. (2012). Evolução convergente em toda a Comunidade na adaptação de insetos a cardenóides tóxicos por substituições na Na,K-ATPase. Proceedings of the National Academy of Sciences, 109(32), 13040–13045. http://doi.org/10.1073/pnas.12021109
- ↑ a b c Yang, L; Ravikanthachari, N; Mariño-Pérez, R; Deshmukh, R; Wu, M; Rosenstein, A; Kunte, K; Song, H; Andolfatto, P. (2019). «Predictability in the evolution of Orthopteran cardenolide insensitivity» (em inglês). Transações filosóficas da Royal Society of London, Series B. 374 (1777): 20180246. doi:10.1098/rstb.2018.0246. PMC 6560278. PMID 31154978.
- ^ Liu, Zhen. Qi, Fei-Yan; Zhou, Xin. Ren, Hai-Qing; Shi, Peng (2014). «Parallel Sites Implicate Functional Convergence of the Hearing Gene Prestin among Echolocating Mammals» (em inglês). Biologia molecular e evolução. 31 (9): 2415–2424.10.1093/molbev/msu194ISSN 1537-1719. PMID 24951728.
- ^ Foote, Andrew D.; Liu, Yue; Thomas, Gregg W. C.; Vinař, Tomáš; Alföldi, Jessica; Deng, Jixin; Dugan, Shannon; Elk, Cornelis E. van; Hunter, Margaret E. (março de 2015). «Convergent evolution of the gennomes of marine mammals» (em inglês). Genética da Natureza. 47 (3): 272–275. doi:10.1038/ng.3198. PMC 46447. PMID 25621460.
- ^ Hu, Yibo. Wu, Qi; Mãe... Shuai. Ma, Tianxiao; Shan, Lei; Wang, Xiao; Nie, Yonggang; Ning, Zemin; Yan, Li (janeiro de 2017). «Comparative genomics reveals convergent evolution between the bamboo-eating gigante and red pandas» (em inglês). Actividades da Academia Nacional de Ciências dos Estados Unidos da América. 114 (5): 1081–1086. Bibcode:2017PNAS..114.1081H. doi:10.1073/pnas.1613870114. PMC 5293045. PMID 28096377.
- ^ Feigin, Charles Y.; Newton, Axel H.; Doronina, Liliya; Schmitz, Jürgen; Hipsley, Christy A.; Mitchell, Kieren J.; Gower, Graham; Llamas, Bastien; Soubrier, Julien (janeiro de 2018). «Genome of the Tasmanian tiger provides insights into the evolution and demography of an extinção marsupial carnivore» (em inglês). Ecologia da Natureza e Evolução. 2 (1): 182–192. doi:10.1038/s41559-017-0417-y. PMID 29230027.
- ^ Partha, Raghavendran; Chauhan, Bharesh K; Ferreira, Zelia; Robinson, Joseph D; Lathrop, Kira; Nischal, Ken K.; Chikina, Maria; Clark, Nathan L. (outubro de 2017). «Subterranean mammals show convergent regressão em genes oculares e potenciadores, juntamente com a adaptação ao túnel». A vida. 6. - Sim.10.7554/eLife.25884. PMC 56430. PMID 29035697.
- ^ Sackton, TB; Grayson, P; Cloutier, A; Hu, Z; Liu, JS; Wheeler, NE; Gardner, PP; Clarke, JA; Baker, AJ; Clamp, M; Edwards, SV (5 de abril de 2019). «Convergent Regulation Evolution and loss of flight in paleognathous Birds» (em inglês). Ciência. 364 (6435): 74–78.10.1126/science.aat7244. PMID 30948549. S2CID 96435050.
- ^ "Como as analogias evoluem?". Universidade da Califórnia Berkeley. Arquivado do original em 2 de abril de 2017. Retrieved 26 de Janeiro 2017.
- ^ Selden, Paul; Nudds, John (2012). Evolução dos Ecossistemas Fosil (2a ed.). CRC Press. p. 133. ISBN 978-1-84076-623-3. Arquivado do original em 15 de fevereiro de 2017. Retrieved 26 de Janeiro 2017.
- ^ Helm, R. R. (18 de novembro de 2015). «Meet Phylliroe: the sea slug that looks and swims like a fish» (em inglês). Notícias do Mar Profundo. Arquivado do original em 26 de julho de 2019. Retrieved 26 de Julho 2019.
- ^ Ballance, Lisa (2016). "O Ambiente Marinho como uma Força Seletiva para Formas Marinhas Secundárias" (PDF). UCSD. Arquivado (PDF) do original em 2 de fevereiro de 2017. Retrieved 19 de Setembro 2019.
- ^ Lento, G. M.; Hickson, R. E.; Chambers, G. K.; Penny, D. (1995). «Use of espectro analysis to test hypotheses on the origin of pinnipeds» (em inglês). Biologia molecular e evolução. 12 (1): 28–52.10.1093/oxfordjournals.molbev.a040189. PMID 7877495.
- ^ Werdelin, L. (1986). «Comparison of Skull Shape in Marsupial and Placental Carnivores» (em inglês). Australian Journal of Zoology. 34 (2): 109–117. doi:10.1071/ZO9860109.
- ^ Liu, Yang; Cotton, James A.; Shen, Bin; Han, Xiuqun; Rossiter, Stephen J.; Zhang, Shuyi (1 de janeiro de 2010). «Convergent sequence evolution between echolocating bats and golfinhos» (em inglês). Biologia atual. 20. (2): R53–R54. doi:10.1016/j.cub.2009.11.058. PMID 20129036. S2CID 16117978.
- ^ Lavoué, Sébastien; Miya, Masaki; Arnegard, Matthew E.; Sullivan, John P.; Hopkins, Carl D.; Nishida, Mutsumi (14 de maio de 2012). «Comparable Ages for the Independent Origins of Electrogenesis in African and South American Weakly Electric Fishes» (em inglês). PLOS UM. 7 (5): e36287. Código: 2012PLoSO...736287L. doi:10.1371/journal.pone.0036287. PMC 3351409. PMID 22606250.
- ^ Roberts, M.B.V. (1986). Biologia: A Abordagem Funcional. Nelson Thornes. p. 274. ISBN 978-0-17-448019-8. Arquivado do original em 12 de setembro de 2016.
- ^ Kozmik, Z; Ruzickova, J; Jonasova, K; Matsumoto, Y.; Vopalensky, P.; Kozmikova, I.; Strnad, H.; Kawamura, S.; Piatigorsky, J.; Paces, V.; Vlcek, C. (1 de julho de 2008). «From the Cover: Assembly of the cnidarian câmera-type eye from vertebrate-like components» (em inglês). Procedimentos da Academia Nacional de Ciências. 105 (26): 8989-8993. Bibcode:2008PNAS..105.8989K. - Sim.10.1073/pnas.0800388105. PMC 2449352. PMID 18577593.
- ^ «Plant and Animal Evolution» (em inglês). Universidade de Waikato. Arquivado do original em 18 de março de 2017. Retrieved 10 de Janeiro 2017.
- ^ Ben-Hamo, Miriam; Muñoz-Garcia, Agustí; Larrain, Paloma; Pinshow, Berry; Korine, Carmi; Williams, Joseph B. (junho de 2016). «A composição lipídica cutânea da asa do morcego e membranas da cauda: um caso de evolução convergente com aves». Proc. R. Soc. B. 283 (1833): 20160636. doi:10.1098/rspb.2016.0636. PMC 4936036. PMID 27335420.
- ^ Alexander, David E. (2015). Sobre a Asa: Insetos, Pterossaros, Pássaros, Bats e a Evolução do Voo Animal. Oxford University Press. p. 28. ISBN 978-0-19-999679-7. Arquivado do original em 14 de fevereiro de 2017. Retrieved 21 de Janeiro 2017.
- ^ «Analogy: Squirrels and Sugar Gliders» (em inglês). Universidade da Califórnia Berkeley. Arquivado do original em 27 de janeiro de 2017. Retrieved 10 de Janeiro 2017.
- ^ Herrera, Carlos M. (1992). "Padrão de atividade e biologia térmica de um dia de Hawkmoth (Macroglossum stellatarum) em condições de verão mediterrânicas. Entomologia Ecológica. 17. (1): 52–56. Bibcode:1992EcoEn...17...52H. doi:10.111111/j.1365-2311.1992.tb01038.x. Hdl:10261 / 446. S2CID 85320151.
- ^ Krenn, Harald W.; Plant, John D.; Szucsich, Nikolaus U. (2005). «Mouthparts of flower-visiting insects» (em inglês). Estrutura e Desenvolvimento de Arthropod. 34 (1): 1–40. doi:10.1016/j.asd.2004.10.002.
- ^ Bauder, Julia A.S.; Lieskonig, Nora R.; Krenn, Harald W. (2011). «The extremamente long-tongued Neotropical borboleta Eurybia lycisca (Riodinidae): Proboscis morphology and flower handling» (em inglês). Estrutura e Desenvolvimento de Arthropod. 40 (2): 122–7. doi:10.1016/j.asd.2010.11.002. PMC 3062012. PMID 21115131.
- ^ Wilhelmi, Andreas P.; Krenn, Harald W. (2012). «Elongated bocaparts of nectar-feeding Meloidae (Coleoptera)» (em inglês). Zoológicomorfologia. 131 (4): 325–37. doi:10.1007/s00435-012-0162-3. S2CID 9194699.
- ^ "Quando é um polegar um polegar?". Compreender a Evolução. Arquivado do original em 16 de outubro de 2015. Retrieved 14 de Agosto 2015.
- ↑ a b d e Edwards, M.; et al. (2010). «Association of the OCA2 Polymorphism His615Arg with Melanin Content in East Asian Populations: Another Evidence of Convergent Evolution of Skin Pigmentation» (em inglês). PLOS Genética. 6 (3): e1000867. - Sim.10.1371/journal.pgen.1000867. PMC 282666. PMID 20221248.
- ^ Meyer, W. K.; et al. (2013). «A evolução convergente da pigmentação da íris azul em primatas tomou caminhos moleculares distintos». Jornal Americano de Antropologia Física. 151 (3): 398–407. doi:10.1002/ajpa.22280. PMC 3746105. PMID 23640739.
- ↑ a b Poppenwimer, Tyler; Mayrose, Itay; DeMalach, Niv (dezembro de 2023). «Revisando a biogeografia global de plantas anuais e perenes». Natureza. 624 (7990): 109–114.2304.13101. Código Bíblico:2023Natur.624..109P. doi:10.1038/s41586-023-06644-x. ISSN 1476-4687. PMC 10830411PMID 37938778. S2CID 260332117.
- ^ Friedman, Jannice (2 de novembro de 2020). «The Evolution of Annual and Perennial Plant Life Histories: Ecological Correlates and Genetic Mechanisms» (em inglês). Revisão Anual de Ecologia, Evolução e Sistemáticos. 51 (1): 461–481. doi:10.1146/annurev-ecolsys-110218-024638. ISSN 1543-592X. S2CID 225237602.
- ^ Hjertaas, Ane C.; Preston, Jill C.; Kainulainen, Kent; Humphreys, Aelys M.; Fjellheim, Siri (2023). «Convergent evolution of the annual life history síndrome from perennial ancestors» (em inglês). Fronteiras em Ciências Vegetais. 13. - Sim.10.3389/fpls.2022.1048656. ISSN 1664-462X. PMC 984627. PMID 36684797.
- ^ Boyko, James D.; Hagen, Eric R.; Beaulieu, Jeremy M.; Vasconcelos, Thais (novembro de 2023). «The evolutionary responses of life-history strategy to climate variability in flowering plants» (em inglês). Novo médico. 240 (4): 1587–1600.10.1111/nph.18971. ISSN 0028-646X. PMID 37194450.
- ^ Williams, B. P.; Johnston, I. G.; Covshoff, S.; Hibberd, J. M. (Setembro de 2013). «Phenotypic landscape inference reveals multiple evolutionary paths to C4 photosynthesis» (em inglês). A vida. 2: e00961. - Sim.10.7554/eLife.00961. PMC 3786385. PMID 24082995.
- ^ Osborne, C. P.; Beerling, D. J. (2006). «Nature's green revolution: the notável evolutionary rise of C4 plants» (em inglês). Transações filosóficas da Royal Society B: Ciências Biológicas. 361 (1465): 173–194. doi:10.1098/rstb.2005.1737. PMC 1626541. PMID 16553316.
- ^ Sage, Rowan; Russell Monson (1999). "16". Biologia da planta C4. Elsevier. pp. 551–580. ISBN 978-0-12-614440-6.
- ^ Zhu, X. G.; Long, S. P.; Ort, D. R. (2008). "Qual é a máxima eficiência com a qual a fotossíntese pode converter a energia solar em biomassa?". Opinião atual em Biotecnologia. 19 (2): 153-159. doi:10.1016/j.copbio.2008.02.004. PMID 18374559. Arquivado do original em 1 de abril de 2019. Retrieved 29 de Dezembro 2018.
- ^ Sage, Rowan; Russell Monson (1999). "7". Biologia da planta C4. Elsevier. pp. 228–229. ISBN 978-0-12-614440-6.
- ^ Kadereit, G.; Borsch, T.; Weising, K.; Freitag, H (2003). "Phylogeny of Amaranthaceae and Chenopodiaceae and the Evolution of C4 Fotossíntese". Jornal Internacional de Ciências Vegetais. 164 (6): 959–86. doi:10.1086/378649. S2CID 83564261.
- ^ Irlanda, Hilary, S.; et al. (2013). «Apple SEPALLATA1/2 -like genes control fruit meat development and ripening» (em inglês). O Jornal Planta. 73 (6): 1044–1056.10.1111/tpj.12094. PMID 23236986.
{{cite journal}}: CS1 maint: vários nomes: lista de autores (link) - ^ Heuvelink, Ep (2005). Tomates. CABI. p. 72. ISBN 978-1-84593-149-0. Arquivado do original em 1 de abril de 2019. Retrieved 17 de Dezembro 2016.
- ^ Lorts, C.; Briggeman, T.; Sang, T. (2008). "Evolução de tipos de frutas e dispersão de sementes: um instantâneo filogenético e ecológico" (PDF). Journal of Systematics and Evolution. 46. (3): 396–404. doi:10.3724/SP.J.1002.2008.08039 (inativo 31 de janeiro de 2024). Arquivado do original (PDF) em 18 de julho de 2013.
{{cite journal}}: Principal CS1: DOI inativo a partir de janeiro 2024 (link) - ^ Lengyel, S.; Gove, A. D.; Latimer, A. M.; Majer, J. D.; Dunn, R. R. (2010). «Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: a global survey» (em inglês). Perspectivas em Ecologia Vegetal, Evolução e Sistemáticos. 12: 43–55. doi:10.1016/j.ppees.2009.08.001.
- ^ Fukushima, K; Fang, X; et al. (2017). «Genome of the pitcher plant Cephalotus reveals genetic changes associated with carnivory» (em inglês). Ecologia da Natureza e Evolução. 1 (3): 0059. Bibcode:2017NatEE...1...59F. doi:10.1038/s41559-016-0059. PMID 28812732.
- ↑ a b c Stayton, C. Tristan (2015). "A definição, reconhecimento e interpretação da evolução convergente, e duas novas medidas para quantificar e avaliar o significado da convergência". Evolução. 69 (8): 2140–2153. doi:10.1111/evo.12729. PMID 26177938. S2CID 3161530.
- ^ Stayton, C. Tristan (2008). "A convergência é surpreendente? Um exame da frequência de convergência em conjuntos de dados simulados". Revista de Biologia Teórica. 252 (1): 1–14. Bibcode:2008JThBi.252....1S. doi:10.1016/j.jtbi.2008.01.008. PMID 18321532.
- ^ Muschick, Moritz; Indermaur, Adrian; Salzburger, Walter (2012). «Convergent Evolution inside an Adaptive Radiation of Cichlid Fishes» (em inglês). Biologia atual. 22 (24): 2362–2368.10.1016/j.cub.2012.10.048. PMID 23159601.
- ^ Arbuckle, Kevin; Bennett, Cheryl M.; Speed, Michael P. (julho de 2014). «Uma medida simples da força da evolução convergente». Métodos em Ecologia e Evolução. 5 (7): 685–693. Bibcode:2014MEcEv...5..685A. doi:10.1111/2041-210X.12195.
- ^ Ingram, Travis; Mahler, D. Luke (1 de maio de 2013). «SURFACE: detecting convergent evolution from comparativa data by setting Ornstein-Uhlenbeck models with stepwise Akaike Information Criterion» (em inglês). Métodos em Ecologia e Evolução. 4 (5): 416–425. Bibcode:2013MEcEv...4.416I. doi:10.1111/2041-210X.12034. S2CID 86382470.
Leitura adicional
- Losos, Jonathan B. (2017). Destinos Improbíveis: Fato, Chance e o Futuro da Evolução. Riverhead Books. ISBN 978-0399184925.
Ligações externas
 Mídia relacionada à evolução Convergent no Wikimedia Commons
Mídia relacionada à evolução Convergent no Wikimedia Commons
| Bases de dados de controle de autoridade: Nacional |
|
|---|