
Botânica

Botânica, também chamada de ciência vegetal(s), biologia vegetal ou fitologia, é a ciência da vida vegetal e um ramo da biologia. Um botânico, cientista de plantas ou fitólogo é um cientista especializado neste campo. O termo "botânica" vem da palavra grega antiga βοτάνη (botanē) que significa "pasto", "ervas" "grama", ou "forragem"; βοτάνη é, por sua vez, derivado de βόσκειν (boskein), "para alimentar" ou "para pastar". Tradicionalmente, a botânica também incluiu o estudo de fungos e algas por micologistas e ficólogos, respectivamente, com o estudo desses três grupos de organismos permanecendo na esfera de interesse do Congresso Internacional de Botânica. Atualmente, os botânicos (no sentido estrito) estudam aproximadamente 410.000 espécies de plantas terrestres, das quais cerca de 391.000 espécies são plantas vasculares (incluindo aproximadamente 369.000 espécies de plantas com flores) e aproximadamente 20.000 são briófitas.
A botânica se originou na pré-história como herbalismo com os esforços dos primeiros humanos para identificar – e depois cultivar – plantas que eram comestíveis, venenosas e possivelmente medicinais, tornando-se um dos primeiros empreendimentos da investigação humana. Os jardins físicos medievais, muitas vezes ligados a mosteiros, continham plantas possivelmente com benefícios medicinais. Foram precursores dos primeiros jardins botânicos ligados a universidades, fundados a partir da década de 1540. Um dos primeiros foi o jardim botânico de Pádua. Esses jardins facilitaram o estudo acadêmico das plantas. Os esforços para catalogar e descrever suas coleções foram os primórdios da taxonomia vegetal e levaram, em 1753, ao sistema binomial de nomenclatura de Carl Linnaeus, que permanece em uso até hoje para nomear todas as espécies biológicas.
Nos séculos 19 e 20, novas técnicas foram desenvolvidas para o estudo das plantas, incluindo métodos de microscopia óptica e imagens de células vivas, microscopia eletrônica, análise do número de cromossomos, química vegetal e estrutura e função de enzimas e outras proteínas. Nas duas últimas décadas do século 20, os botânicos exploraram as técnicas de análise genética molecular, incluindo genômica e proteômica e sequências de DNA para classificar as plantas com mais precisão.
A botânica moderna é um assunto amplo e multidisciplinar com contribuições e insights da maioria das outras áreas da ciência e tecnologia. Os tópicos de pesquisa incluem o estudo da estrutura vegetal, crescimento e diferenciação, reprodução, bioquímica e metabolismo primário, produtos químicos, desenvolvimento, doenças, relações evolutivas, sistemática e taxonomia vegetal. Os temas dominantes na ciência vegetal do século XXI são a genética molecular e a epigenética, que estudam os mecanismos e o controle da expressão gênica durante a diferenciação de células e tecidos vegetais. A pesquisa botânica tem diversas aplicações no fornecimento de alimentos básicos, materiais como madeira, óleo, borracha, fibras e drogas, na moderna horticultura, agricultura e silvicultura, propagação de plantas, reprodução e modificação genética, na síntese de produtos químicos e matérias-primas para construção e produção de energia, na gestão ambiental e na manutenção da biodiversidade.
História
Botânica primitiva

A botânica originou-se como herbalismo, o estudo e uso de plantas por suas possíveis propriedades medicinais. Os primeiros registros da história da botânica incluem muitos escritos antigos e classificações de plantas. Exemplos de trabalhos botânicos antigos foram encontrados em textos antigos da Índia que datam de antes de 1100 aC, Egito Antigo, em escritos arcaicos do Avestan e em trabalhos da China supostamente anteriores a 221 aC.
A botânica moderna tem suas raízes na Grécia Antiga, especificamente em Teofrasto (c. 371–287 aC), um aluno de Aristóteles que inventou e descreveu muitos de seus princípios e é amplamente considerado na comunidade científica como o "Pai da Botânica". Suas principais obras, Investigação das plantas e Sobre as causas das plantas, constituem as contribuições mais importantes para a ciência botânica até a Idade Média, quase dezessete séculos depois.
Outra obra da Grécia Antiga que teve um impacto inicial na botânica é De materia medica, uma enciclopédia de cinco volumes sobre fitoterapia preliminar escrita em meados do primeiro século pelo médico e farmacologista grego Pedanius Dioscórides. De materia medica foi amplamente lido por mais de 1.500 anos. Contribuições importantes do mundo muçulmano medieval incluem a Agricultura Nabateana de Ibn Wahshiyya, o Livro das Plantas de Abū Ḥanīfa Dīnawarī (828–896) e Ibn A classificação dos solos de Bassal. No início do século 13, Abu al-Abbas al-Nabati e Ibn al-Baitar (falecido em 1248) escreveram sobre botânica de maneira sistemática e científica.
Em meados do século XVI, foram fundados jardins botânicos em várias universidades italianas. O jardim botânico de Pádua em 1545 é geralmente considerado o primeiro que ainda está em sua localização original. Esses jardins continuaram o valor prático dos antigos "jardins físicos", frequentemente associados a mosteiros, nos quais as plantas eram cultivadas para usos medicinais suspeitos. Eles apoiaram o crescimento da botânica como disciplina acadêmica. Foram ministradas palestras sobre as plantas cultivadas nos jardins. Os jardins botânicos chegaram muito mais tarde ao norte da Europa; o primeiro na Inglaterra foi o Jardim Botânico da Universidade de Oxford em 1621.
O médico alemão Leonhart Fuchs (1501–1566) foi um dos "três pais alemães da botânica", juntamente com o teólogo Otto Brunfels (1489–1534) e o médico Hieronymus Bock (1498–1554) (também chamado Hieronymus Tragus). Fuchs e Brunfels romperam com a tradição de copiar obras anteriores para fazer suas próprias observações originais. Bock criou seu próprio sistema de classificação de plantas.
O médico Valerius Cordus (1515–1544) escreveu uma Historia Plantarum à base de plantas botânica e farmacologicamente importante em 1544 e uma farmacopeia de importância duradoura, o Dispensatorium em 1546. Naturalista Conrad von Gesner (1516–1565) e o fitoterapeuta John Gerard (1545–c. 1611) publicaram coberturas de ervas os supostos usos medicinais das plantas. O naturalista Ulisse Aldrovandi (1522–1605) foi considerado o pai da história natural, que incluía o estudo das plantas. Em 1665, usando um microscópio antigo, o polímata Robert Hooke descobriu células, um termo que ele cunhou, em cortiça e, pouco tempo depois, em tecido vegetal vivo.
Botânica do início da era moderna

Durante o século XVIII, foram desenvolvidos sistemas de identificação de plantas comparáveis a chaves dicotômicas, onde plantas não identificadas são colocadas em grupos taxonômicos (por exemplo, família, gênero e espécie) fazendo uma série de escolhas entre pares de caracteres. A escolha e a sequência dos caracteres podem ser artificiais em chaves destinadas exclusivamente à identificação (chaves diagnósticas) ou mais próximas da ordem natural ou filética dos táxons em chaves sinóticas. No século 18, novas plantas para estudo chegavam à Europa em números crescentes de países recém-descobertos e das colônias européias em todo o mundo. Em 1753, Carl von Linné (Carl Linnaeus) publicou seu Species Plantarum, uma classificação hierárquica de espécies de plantas que continua sendo o ponto de referência para a nomenclatura botânica moderna. Isso estabeleceu um esquema de nomenclatura binomial ou de duas partes padronizado, onde o primeiro nome representava o gênero e o segundo identificava as espécies dentro do gênero. Para fins de identificação, o Systema Sexuale de Linnaeus classificou as plantas em 24 grupos de acordo com o número de seus órgãos sexuais masculinos. O 24º grupo, Cryptogamia, incluiu todas as plantas com partes reprodutivas ocultas, musgos, hepáticas, samambaias, algas e fungos.
O crescente conhecimento da anatomia, morfologia e ciclos de vida das plantas levou à percepção de que havia mais afinidades naturais entre as plantas do que o sistema sexual artificial de Linnaeus. Adanson (1763), de Jussieu (1789) e Candolle (1819) propuseram vários sistemas naturais alternativos de classificação que agrupavam plantas usando uma gama mais ampla de caracteres compartilhados e foram amplamente seguidos. O sistema candolliano refletia suas ideias sobre a progressão da complexidade morfológica e o sistema Bentham & O sistema Hooker, influente até meados do século XIX, foi influenciado pela abordagem de Candolle. A publicação de Darwin da Origem das Espécies em 1859 e seu conceito de descendência comum exigiram modificações no sistema Candolliano para refletir as relações evolutivas como distintas da mera similaridade morfológica.
A botânica foi muito estimulada pelo surgimento das primeiras espécies "modernas" livro didático, Grundzüge der Wissenschaftlichen Botanik de Matthias Schleiden, publicado em inglês em 1849 como Princípios de Botânica Científica. Schleiden foi um microscopista e um dos primeiros anatomistas de plantas que co-fundou a teoria celular com Theodor Schwann e Rudolf Virchow e foi um dos primeiros a entender o significado do núcleo celular que havia sido descrito por Robert Brown em 1831. Em 1855, Adolf Fick formulou as leis de Fick que permitiram o cálculo das taxas de difusão molecular em sistemas biológicos.

Botânica moderna tardia
Com base na teoria do gene-cromossomo da hereditariedade que se originou com Gregor Mendel (1822–1884), August Weismann (1834–1914) provou que a herança ocorre apenas por meio de gametas. Nenhuma outra célula pode transmitir caracteres herdados. O trabalho de Katherine Esau (1898–1997) sobre anatomia vegetal ainda é um dos principais fundamentos da botânica moderna. Seus livros Plant Anatomy e Anatomy of Seed Plants têm sido os principais textos de biologia estrutural de plantas por mais de meio século.

A disciplina de ecologia vegetal foi iniciada no final do século 19 por botânicos como Eugenius Warming, que produziu a hipótese de que as plantas formam comunidades, e seu mentor e sucessor Christen C. Raunkiær, cujo sistema para descrever formas de vida vegetal ainda está em usar hoje. O conceito de que a composição de comunidades de plantas, como florestas temperadas de folhas largas, muda por um processo de sucessão ecológica foi desenvolvido por Henry Chandler Cowles, Arthur Tansley e Frederic Clements. Clements é creditado com a ideia de vegetação clímax como a vegetação mais complexa que um ambiente pode suportar e Tansley introduziu o conceito de ecossistemas na biologia. Com base no extenso trabalho anterior de Alphonse de Candolle, Nikolai Vavilov (1887-1943) produziu relatos da biogeografia, centros de origem e história evolutiva de plantas econômicas.
Particularmente desde meados da década de 1960, houve avanços na compreensão da física dos processos fisiológicos das plantas, como a transpiração (o transporte de água dentro dos tecidos vegetais), a dependência da temperatura das taxas de evaporação da água da superfície da folha e a estrutura molecular difusão de vapor de água e dióxido de carbono através das aberturas estomáticas. Esses desenvolvimentos, juntamente com novos métodos para medir o tamanho das aberturas estomáticas e a taxa de fotossíntese, permitiram a descrição precisa das taxas de troca gasosa entre as plantas e a atmosfera. Inovações em análise estatística por Ronald Fisher, Frank Yates e outros na Rothamsted Experimental Station facilitaram o planejamento experimental racional e a análise de dados na pesquisa botânica. A descoberta e identificação dos hormônios vegetais auxina por Kenneth V. Thimann em 1948 permitiu a regulação do crescimento vegetal por produtos químicos aplicados externamente. Frederick Campion Steward foi pioneiro em técnicas de micropropagação e cultura de tecidos vegetais controlada por hormônios vegetais. A auxina sintética ácido 2,4-diclorofenoxiacético ou 2,4-D foi um dos primeiros herbicidas sintéticos comerciais.

Os desenvolvimentos do século 20 na bioquímica vegetal foram impulsionados por técnicas modernas de análise química orgânica, como espectroscopia, cromatografia e eletroforese. Com o surgimento das abordagens biológicas relacionadas à escala molecular de biologia molecular, genômica, proteômica e metabolômica, a relação entre o genoma da planta e a maioria dos aspectos da bioquímica, fisiologia, morfologia e comportamento das plantas pode ser submetida a análises experimentais detalhadas. O conceito originalmente declarado por Gottlieb Haberlandt em 1902 de que todas as células vegetais são totipotentes e podem ser cultivadas in vitro permitiu o uso da engenharia genética experimentalmente para eliminar um gene ou genes responsáveis por uma característica específica, ou para adicionar genes como GFP que relatam quando um gene de interesse está sendo expresso. Essas tecnologias permitem o uso biotecnológico de plantas inteiras ou culturas de células vegetais cultivadas em biorreatores para sintetizar pesticidas, antibióticos ou outros produtos farmacêuticos, bem como a aplicação prática de culturas geneticamente modificadas projetadas para características como melhor rendimento.
A morfologia moderna reconhece um continuum entre as principais categorias morfológicas de raiz, caule (cauloma), folha (filoma) e tricoma. Além disso, enfatiza a dinâmica estrutural. A sistemática moderna visa refletir e descobrir as relações filogenéticas entre as plantas. A filogenética molecular moderna ignora em grande parte os caracteres morfológicos, baseando-se nas sequências de DNA como dados. A análise molecular das sequências de DNA da maioria das famílias de plantas com flores permitiu que o Angiosperm Phylogeny Group publicasse em 1998 uma filogenia de plantas com flores, respondendo a muitas das questões sobre as relações entre famílias e espécies de angiospermas. A possibilidade teórica de um método prático para identificação de espécies de plantas e variedades comerciais por DNA barcoding é objeto de intensa pesquisa atual.
Escopo e importância

O estudo das plantas é vital porque elas sustentam quase toda a vida animal na Terra, gerando uma grande proporção de oxigênio e alimentos que fornecem aos humanos e outros organismos a respiração aeróbica com a energia química de que precisam para existir. Plantas, algas e cianobactérias são os principais grupos de organismos que realizam a fotossíntese, processo que utiliza a energia da luz solar para converter água e dióxido de carbono em açúcares que podem ser utilizados tanto como fonte de energia química quanto de moléculas orgânicas que são utilizadas nos componentes estruturais das células. Como subproduto da fotossíntese, as plantas liberam oxigênio na atmosfera, um gás necessário para quase todos os seres vivos realizarem a respiração celular. Além disso, eles são influentes nos ciclos globais de carbono e água e as raízes das plantas ligam e estabilizam os solos, evitando a erosão do solo. As plantas são cruciais para o futuro da sociedade humana, pois fornecem alimentos, oxigênio, bioquímicos e produtos para as pessoas, além de criar e preservar o solo.
Historicamente, todos os seres vivos foram classificados como animais ou plantas e a botânica abrangeu o estudo de todos os organismos não considerados animais. Os botânicos examinam as funções internas e os processos dentro das organelas, células, tecidos, plantas inteiras, populações de plantas e comunidades de plantas. Em cada um desses níveis, um botânico pode se preocupar com a classificação (taxonomia), filogenia e evolução, estrutura (anatomia e morfologia) ou função (fisiologia) da vida vegetal.
A definição mais estrita de "planta" inclui apenas as "plantas terrestres" ou embriófitos, que incluem plantas com sementes (gimnospermas, incluindo os pinheiros e plantas com flores) e os criptógamos de esporos livres, incluindo samambaias, musgos, hepáticas, antóceros e musgos. Os embriófitos são eucariotos multicelulares descendentes de um ancestral que obteve sua energia da luz solar pela fotossíntese. Eles têm ciclos de vida com fases haploides e diploides alternadas. A fase sexual haploide dos embriófitos, conhecida como gametófito, nutre o esporófito do embrião diplóide em desenvolvimento dentro de seus tecidos por pelo menos parte de sua vida, mesmo nas plantas com sementes, onde o próprio gametófito é nutrido por seu esporófito parental. Outros grupos de organismos que foram previamente estudados por botânicos incluem bactérias (agora estudadas em bacteriologia), fungos (micologia) – incluindo fungos formadores de líquen (liquenologia), algas não clorofíticas (ficologia) e vírus (virologia). No entanto, a atenção ainda é dada a esses grupos por botânicos, e fungos (incluindo líquens) e protistas fotossintéticos são geralmente abordados em cursos introdutórios de botânica.
Os paleobotânicos estudam plantas antigas no registro fóssil para fornecer informações sobre a história evolutiva das plantas. Acredita-se que as cianobactérias, os primeiros organismos fotossintéticos liberadores de oxigênio na Terra, tenham dado origem ao ancestral das plantas ao entrar em uma relação endossimbiótica com um eucarioto primitivo, tornando-se finalmente os cloroplastos nas células vegetais. As novas plantas fotossintéticas (juntamente com suas parentes algas) aceleraram o aumento do oxigênio atmosférico iniciado pelas cianobactérias, mudando a antiga atmosfera reduzida e livre de oxigênio para uma em que o oxigênio livre é abundante há mais de 2 bilhões de anos.
Entre as questões botânicas importantes do século 21 estão o papel das plantas como produtores primários no ciclo global dos ingredientes básicos da vida: energia, carbono, oxigênio, nitrogênio e água, e maneiras pelas quais nossa administração de plantas pode ajudar a abordar as questões ambientais globais de gestão de recursos, conservação, segurança alimentar humana, organismos biologicamente invasivos, sequestro de carbono, mudança climática e sustentabilidade.
Nutrição humana

Praticamente todos os alimentos básicos vêm diretamente da produção primária das plantas ou indiretamente dos animais que os comem. As plantas e outros organismos fotossintéticos estão na base da maioria das cadeias alimentares porque utilizam a energia do sol e os nutrientes do solo e da atmosfera, convertendo-os numa forma que pode ser utilizada pelos animais. Isso é o que os ecologistas chamam de primeiro nível trófico. As formas modernas dos principais alimentos básicos, como cânhamo, teff, milho, arroz, trigo e outras gramíneas de cereais, leguminosas, bananas e bananas-da-terra, bem como cânhamo, linho e algodão cultivados por suas fibras, são o resultado da seleção pré-histórica ao longo de milhares de anos entre plantas ancestrais silvestres com as características mais desejáveis.
Os botânicos estudam como as plantas produzem alimentos e como aumentar os rendimentos, por exemplo, por meio do melhoramento de plantas, tornando seu trabalho importante para a capacidade da humanidade de alimentar o mundo e fornecer segurança alimentar para as gerações futuras. Os botânicos também estudam ervas daninhas, que são um problema considerável na agricultura, e a biologia e controle de patógenos de plantas na agricultura e ecossistemas naturais. Etnobotânica é o estudo das relações entre as plantas e as pessoas. Quando aplicada à investigação das relações históricas entre plantas e pessoas, a etnobotânica pode ser referida como arqueobotânica ou paleoetnobotânica. Algumas das primeiras relações planta-pessoas surgiram entre os povos indígenas do Canadá na identificação de plantas comestíveis de plantas não comestíveis. Essa relação dos indígenas com as plantas foi registrada por etnobotânicos.
Bioquímica vegetal
A bioquímica vegetal é o estudo dos processos químicos usados pelas plantas. Alguns desses processos são usados em seu metabolismo primário, como o ciclo fotossintético de Calvin e o metabolismo do ácido crassuláceo. Outros fabricam materiais especializados, como celulose e lignina, usados para construir seus corpos, e produtos secundários, como resinas e compostos aromáticos.

Plantas e vários outros grupos de eucariontes fotossintéticos conhecidos coletivamente como "algas" têm organelas únicas conhecidas como cloroplastos. Acredita-se que os cloroplastos sejam descendentes de cianobactérias que formaram relações endossimbióticas com ancestrais de plantas e algas. Cloroplastos e cianobactérias contêm o pigmento verde-azulado clorofila a. A clorofila a (assim como sua prima clorofila b específica para plantas e algas verdes) absorve a luz nas partes azul-violeta e laranja/vermelha do espectro enquanto reflete e transmite a luz verde que vemos como a cor característica desses organismos. A energia na luz vermelha e azul que esses pigmentos absorvem é usada pelos cloroplastos para produzir compostos de carbono ricos em energia a partir de dióxido de carbono e água pela fotossíntese oxigênica, um processo que gera oxigênio molecular (O2) como um subproduto.

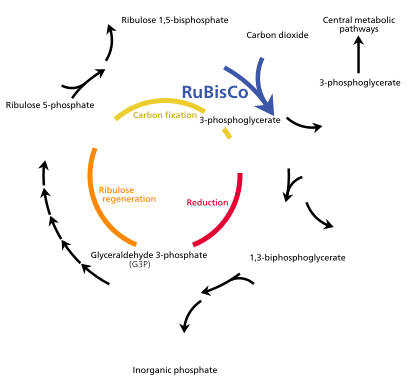


A energia da luz capturada pela clorofila a está inicialmente na forma de elétrons (e depois um gradiente de prótons) que é usado para fazer moléculas de ATP e NADPH que temporariamente armazenam e transportam energia. Sua energia é usada nas reações independentes de luz do ciclo de Calvin pela enzima rubisco para produzir moléculas do açúcar de 3 carbonos gliceraldeído 3-fosfato (G3P). O gliceraldeído 3-fosfato é o primeiro produto da fotossíntese e a matéria-prima a partir da qual a glicose e quase todas as outras moléculas orgânicas de origem biológica são sintetizadas. Parte da glicose é convertida em amido, que é armazenado no cloroplasto. O amido é o armazenamento de energia característico da maioria das plantas terrestres e algas, enquanto a inulina, um polímero de frutose, é usado para o mesmo propósito na família de girassóis Asteraceae. Parte da glicose é convertida em sacarose (açúcar de mesa comum) para exportação para o restante da planta.
Ao contrário dos animais (que não possuem cloroplastos), as plantas e seus parentes eucariontes delegaram muitos papéis bioquímicos aos seus cloroplastos, incluindo a síntese de todos os seus ácidos graxos e da maioria dos aminoácidos. Os ácidos graxos que os cloroplastos produzem são usados para muitas coisas, como fornecer material para construir membranas celulares e produzir o polímero cutina encontrado na cutícula da planta que protege as plantas terrestres de secar.
As plantas sintetizam vários polímeros únicos, como as moléculas de polissacarídeos celulose, pectina e xiloglucana, a partir das quais a parede celular da planta terrestre é construída. As plantas terrestres vasculares produzem lignina, um polímero usado para fortalecer as paredes celulares secundárias dos traqueídeos e vasos do xilema para evitar que entrem em colapso quando uma planta suga água através deles sob estresse hídrico. A lignina também é usada em outros tipos de células, como fibras de esclerênquima, que fornecem suporte estrutural para uma planta e é um dos principais constituintes da madeira. A esporopolenina é um polímero quimicamente resistente encontrado nas paredes celulares externas de esporos e pólen de plantas terrestres responsáveis pela sobrevivência dos primeiros esporos de plantas terrestres e do pólen de plantas com sementes no registro fóssil. É amplamente considerado como um marcador para o início da evolução das plantas terrestres durante o período Ordoviciano. A concentração de dióxido de carbono na atmosfera hoje é muito menor do que era quando as plantas surgiram na terra durante os períodos Ordoviciano e Siluriano. Muitas monocotiledôneas, como milho e abacaxi, e algumas dicotiledôneas, como as Asteraceae, evoluíram independentemente, como o metabolismo do ácido crassuláceo e a via de fixação de carbono C4 para a fotossíntese, que evitam as perdas resultantes da fotorrespiração na via mais comum de fixação de carbono C3. Essas estratégias bioquímicas são exclusivas das plantas terrestres.
Remédios e materiais
A fitoquímica é um ramo da bioquímica vegetal que se preocupa principalmente com as substâncias químicas produzidas pelas plantas durante o metabolismo secundário. Alguns desses compostos são toxinas, como o alcaloide coniina da cicuta. Outros, como os óleos essenciais óleo de hortelã-pimenta e óleo de limão, são úteis por seu aroma, como aromatizantes e especiarias (por exemplo, capsaicina) e na medicina como produtos farmacêuticos como no ópio das papoulas. Muitas drogas medicinais e recreativas, como o tetrahidrocanabinol (ingrediente ativo da cannabis), a cafeína, a morfina e a nicotina vêm diretamente das plantas. Outros são simples derivados de produtos naturais botânicos. Por exemplo, o analgésico aspirina é o éster acetil do ácido salicílico, originalmente isolado da casca dos salgueiros, e uma ampla variedade de analgésicos opiáceos, como a heroína, são obtidos pela modificação química da morfina obtida da papoula do ópio. Estimulantes populares vêm de plantas, como a cafeína do café, chá e chocolate, e a nicotina do tabaco. A maioria das bebidas alcoólicas vem da fermentação de produtos vegetais ricos em carboidratos, como cevada (cerveja), arroz (sake) e uvas (vinho). Os nativos americanos usaram várias plantas como forma de tratar doenças ou enfermidades por milhares de anos. Esse conhecimento que os nativos americanos têm sobre as plantas foi registrado por etnobotânicos e, por sua vez, usado por empresas farmacêuticas como uma forma de descoberta de medicamentos.
As plantas podem sintetizar corantes e pigmentos coloridos, como as antocianinas responsáveis pela cor vermelha do vinho tinto, amarelo soldado e azul, usados juntos para produzir o verde Lincoln, indoxil, fonte do corante azul índigo tradicionalmente usado para tingir denim e o pigmentos artísticos gamboge e rose madder.
Açúcar, amido, algodão, linho, cânhamo, alguns tipos de corda, madeira e painéis de partículas, papiro e papel, óleos vegetais, cera e borracha natural são exemplos de materiais comercialmente importantes feitos de tecidos vegetais ou seus produtos secundários. O carvão, uma forma pura de carbono feita por pirólise de madeira, tem uma longa história como combustível para fundição de metais, como material filtrante e adsorvente e como material artístico e é um dos três ingredientes da pólvora. A celulose, o polímero orgânico mais abundante do mundo, pode ser convertida em energia, combustíveis, materiais e matéria-prima química. Os produtos feitos de celulose incluem rayon e celofane, pasta de papel de parede, biobutanol e algodão para armas. A cana-de-açúcar, a colza e a soja são algumas das plantas com alto teor de açúcar ou óleo fermentável que são utilizadas como fontes de biocombustíveis, importantes alternativas aos combustíveis fósseis, como o biodiesel. Sweetgrass foi usado pelos nativos americanos para afastar insetos como mosquitos. Essas propriedades repelentes de insetos da erva-doce foram posteriormente encontradas pela American Chemical Society nas moléculas fitol e cumarina.
Ecologia vegetal

A ecologia vegetal é a ciência das relações funcionais entre as plantas e seus habitats - os ambientes onde elas completam seus ciclos de vida. Os ecologistas de plantas estudam a composição das floras locais e regionais, sua biodiversidade, diversidade genética e aptidão, a adaptação das plantas ao seu ambiente e suas interações competitivas ou mutualísticas com outras espécies. Alguns ecologistas até contam com dados empíricos de povos indígenas coletados por etnobotânicos. Esta informação pode transmitir uma grande quantidade de informações sobre como a terra era há milhares de anos e como ela mudou ao longo desse tempo. Os objetivos da ecologia vegetal são entender as causas de seus padrões de distribuição, produtividade, impacto ambiental, evolução e respostas às mudanças ambientais.
As plantas dependem de certos fatores edáficos (solo) e climáticos em seu ambiente, mas também podem modificar esses fatores. Por exemplo, eles podem alterar o albedo de seu ambiente, aumentar a interceptação do escoamento, estabilizar solos minerais e desenvolver seu conteúdo orgânico e afetar a temperatura local. As plantas competem com outros organismos em seu ecossistema por recursos. Eles interagem com seus vizinhos em uma variedade de escalas espaciais em grupos, populações e comunidades que constituem coletivamente a vegetação. Regiões com tipos de vegetação característicos e plantas dominantes, bem como fatores abióticos e bióticos semelhantes, clima e geografia compõem biomas como tundra ou floresta tropical.
Os herbívoros comem plantas, mas as plantas podem se defender e algumas espécies são parasitas ou mesmo carnívoras. Outros organismos formam relações mutuamente benéficas com as plantas. Por exemplo, fungos micorrízicos e rizóbios fornecem nutrientes às plantas em troca de alimentos, formigas são recrutadas por plantas formigas para fornecer proteção, abelhas, morcegos e outros animais polinizam flores e humanos e outros animais atuam como vetores de dispersão para espalhar esporos e sementes.
Plantas, clima e mudanças ambientais
As respostas das plantas ao clima e outras mudanças ambientais podem informar nossa compreensão de como essas mudanças afetam a função e a produtividade do ecossistema. Por exemplo, a fenologia da planta pode ser um proxy útil para a temperatura na climatologia histórica e o impacto biológico da mudança climática e do aquecimento global. Palinologia, a análise de depósitos de pólen fóssil em sedimentos de milhares ou milhões de anos atrás permite a reconstrução de climas passados. As estimativas das concentrações atmosféricas de CO 2 desde o Paleozóico foram obtidas a partir das densidades estomáticas e das formas e tamanhos das folhas de antigas plantas terrestres. A destruição do ozônio pode expor as plantas a níveis mais altos de radiação ultravioleta B (UV-B), resultando em taxas de crescimento mais baixas. Além disso, informações de estudos de ecologia de comunidades, sistemática de plantas e taxonomia são essenciais para entender a mudança de vegetação, destruição de habitat e extinção de espécies.
Genética

A herança nas plantas segue os mesmos princípios fundamentais da genética que em outros organismos multicelulares. Gregor Mendel descobriu as leis genéticas da herança estudando características herdadas, como a forma em Pisum sativum (ervilhas). O que Mendel aprendeu com o estudo das plantas teve benefícios de longo alcance fora da botânica. Da mesma forma, "genes saltadores" foram descobertos por Barbara McClintock enquanto ela estudava milho. No entanto, existem algumas diferenças genéticas distintas entre plantas e outros organismos.
Os limites de espécies em plantas podem ser mais fracos do que em animais, e híbridos de espécies cruzadas são frequentemente possíveis. Um exemplo familiar é a hortelã-pimenta, Mentha × piperita, um híbrido estéril entre Mentha aquatica e hortelã, Mentha spicata. As muitas variedades cultivadas de trigo são o resultado de múltiplos cruzamentos inter e intraespecíficos entre espécies silvestres e seus híbridos. As angiospermas com flores monóicas geralmente têm mecanismos de auto-incompatibilidade que operam entre o pólen e o estigma, de modo que o pólen falha em alcançar o estigma ou não germina e produz gametas masculinos. Este é um dos vários métodos usados pelas plantas para promover o cruzamento. Em muitas plantas terrestres, os gametas masculino e feminino são produzidos por indivíduos separados. Essas espécies são ditas dióicas quando se referem a esporófitos de plantas vasculares e dióicas quando se referem a gametófitos briófitos.
Ao contrário dos animais superiores, onde a partenogênese é rara, a reprodução assexuada pode ocorrer nas plantas por vários mecanismos diferentes. A formação de tubérculos caulinares na batata é um exemplo. Particularmente em habitats árticos ou alpinos, onde as oportunidades de fertilização de flores por animais são raras, plântulas ou bulbos podem se desenvolver em vez de flores, substituindo a reprodução sexual por reprodução assexuada e dando origem a populações clonais geneticamente idênticas ao pai. Este é um dos vários tipos de apomixia que ocorrem nas plantas. A apomixia também pode acontecer em uma semente, produzindo uma semente que contém um embrião geneticamente idêntico ao pai.
A maioria dos organismos que se reproduzem sexualmente são diplóides, com cromossomos pareados, mas a duplicação do número de cromossomos pode ocorrer devido a erros na citocinese. Isso pode ocorrer no início do desenvolvimento para produzir um organismo autopoliploide ou parcialmente autopoliploide, ou durante processos normais de diferenciação celular para produzir alguns tipos de células que são poliploides (endopoliploidia) ou durante a formação de gametas. Uma planta alopoliplóide pode resultar de um evento de hibridação entre duas espécies diferentes. Tanto as plantas autopoliplóides quanto as alopoliploides geralmente podem se reproduzir normalmente, mas podem ser incapazes de cruzar com sucesso com a população parental porque há uma incompatibilidade no número de cromossomos. Essas plantas que são reprodutivamente isoladas das espécies progenitoras, mas vivem na mesma área geográfica, podem ter sucesso suficiente para formar uma nova espécie. Alguns poliplóides de plantas estéreis ainda podem se reproduzir vegetativamente ou por apomixia de sementes, formando populações clonais de indivíduos idênticos. O trigo duro é um alopoliploide tetraplóide fértil, enquanto o trigo pão é um hexaploide fértil. A banana comercial é um exemplo de um híbrido triploide estéril e sem sementes. O dente-de-leão comum é um triplóide que produz sementes viáveis por sementes apomíticas.
Como em outros eucariotos, a herança de organelas endossimbióticas como mitocôndrias e cloroplastos em plantas não é mendeliana. Os cloroplastos são herdados através do progenitor masculino nas gimnospermas, mas frequentemente através do progenitor feminino nas plantas com flores.
Genética molecular

Uma quantidade considerável de novos conhecimentos sobre a função da planta vem de estudos da genética molecular de plantas modelo, como o agrião Thale, Arabidopsis thaliana, uma espécie de erva daninha da família da mostarda (Brassicaceae). O genoma ou informação hereditária contida nos genes dessa espécie é codificado por cerca de 135 milhões de pares de bases de DNA, formando um dos menores genomas entre as plantas com flores. A Arabidopsis foi a primeira planta a ter seu genoma sequenciado, em 2000. O sequenciamento de alguns outros genomas relativamente pequenos, de arroz (Oryza sativa) e Brachypodium distachyon, tornou-os importantes espécies modelo para a compreensão da genética, biologia celular e molecular de cereais, gramíneas e monocotiledôneas em geral.
Plantas modelo como Arabidopsis thaliana são usadas para estudar a biologia molecular das células vegetais e do cloroplasto. Idealmente, esses organismos têm genomas pequenos, bem conhecidos ou completamente sequenciados, pequena estatura e tempos de geração curtos. O milho tem sido usado para estudar mecanismos de fotossíntese e carregamento de açúcar no floema em plantas C4. A alga verde unicelular Chlamydomonas reinhardtii, embora não seja um embriófito em si, contém um cloroplasto de pigmentação verde semelhante ao das plantas terrestres, tornando-o útil para estudo. Uma alga vermelha Cyanidioschyzon merolae também foi usada para estudar algumas funções básicas do cloroplasto. Espinafre, ervilha, soja e musgo Physcomitrella patens são comumente usados para estudar a biologia celular de plantas.
Agrobacterium tumefaciens, uma bactéria da rizosfera do solo, pode se ligar a células vegetais e infectá-las com um plasmídeo Ti indutor de calo por transferência horizontal de genes, causando uma infecção de calo chamada doença da galha da coroa. Schell e Van Montagu (1977) levantaram a hipótese de que o plasmídeo Ti poderia ser um vetor natural para a introdução do gene Nif responsável pela fixação de nitrogênio nos nódulos radiculares de leguminosas e outras espécies vegetais. Hoje, a modificação genética do plasmídeo Ti é uma das principais técnicas para introdução de transgenes em plantas e criação de cultivos geneticamente modificados.
Epigenética
A epigenética é o estudo das mudanças hereditárias na função do gene que não podem ser explicadas por mudanças na sequência de DNA subjacente, mas fazem com que os genes do organismo se comportem (ou "se expressem") de maneira diferente. Um exemplo de mudança epigenética é a marcação dos genes pela metilação do DNA, que determina se eles serão expressos ou não. A expressão gênica também pode ser controlada por proteínas repressoras que se ligam a regiões silenciadoras do DNA e impedem que aquela região do código de DNA seja expressa. Marcas epigenéticas podem ser adicionadas ou removidas do DNA durante fases programadas do desenvolvimento da planta, e são responsáveis, por exemplo, pelas diferenças entre anteras, pétalas e folhas normais, apesar de todas terem o mesmo código genético subjacente. Mudanças epigenéticas podem ser temporárias ou podem permanecer através de sucessivas divisões celulares pelo restante da vida da célula. Algumas alterações epigenéticas demonstraram ser hereditárias, enquanto outras são redefinidas nas células germinativas.
Mudanças epigenéticas na biologia eucariótica servem para regular o processo de diferenciação celular. Durante a morfogênese, as células-tronco totipotentes tornam-se as várias linhas celulares pluripotentes do embrião, que por sua vez se tornam células totalmente diferenciadas. Um único óvulo fertilizado, o zigoto, dá origem a muitos tipos diferentes de células vegetais, incluindo parênquima, elementos de vasos do xilema, tubos crivados do floema, células protetoras da epiderme, etc., à medida que continua a se dividir. O processo resulta da ativação epigenética de alguns genes e inibição de outros.
Ao contrário dos animais, muitas células vegetais, particularmente as do parênquima, não se diferenciam terminalmente, permanecendo totipotentes com a capacidade de dar origem a uma nova planta individual. As exceções incluem células altamente lignificadas, o esclerênquima e o xilema que estão mortos na maturidade e os tubos crivados do floema que carecem de núcleos. Embora as plantas usem muitos dos mesmos mecanismos epigenéticos dos animais, como a remodelação da cromatina, uma hipótese alternativa é que as plantas definem seus padrões de expressão gênica usando informações posicionais do ambiente e das células circundantes para determinar seu destino de desenvolvimento.
Mudanças epigenéticas podem levar a paramutações, que não seguem as regras da herança mendeliana. Essas marcas epigenéticas são transmitidas de uma geração para outra, com um alelo induzindo uma mudança no outro.
Evolução vegetal

Os cloroplastos das plantas têm várias semelhanças bioquímicas, estruturais e genéticas com as cianobactérias (comumente, mas incorretamente conhecidas como "algas verde-azuladas") e acredita-se que sejam derivadas de uma antiga relação endossimbiótica entre uma célula eucariótica ancestral e uma cianobactéria residente.
As algas são um grupo polifilético e estão colocadas em várias divisões, algumas mais relacionadas com as plantas do que outras. Existem muitas diferenças entre eles em características como composição da parede celular, bioquímica, pigmentação, estrutura do cloroplasto e reservas de nutrientes. A divisão de algas Charophyta, irmã da divisão de algas verdes Chlorophyta, é considerada o ancestral das plantas verdadeiras. A classe Charophyte Charophyceae e o sub-reino das plantas terrestres Embryophyta juntos formam o grupo monofilético ou clado Streptophytina.
As plantas terrestres não vasculares são embriófitas que não possuem os tecidos vasculares xilema e floema. Eles incluem musgos, hepáticas e antóceros. Plantas vasculares pteridofíticas com xilema e floema verdadeiros que se reproduzem por esporos germinando em gametófitos de vida livre evoluíram durante o período Siluriano e se diversificaram em várias linhagens durante o final do Siluriano e início do Devoniano. Representantes dos licopódios sobreviveram até os dias atuais. No final do período Devoniano, vários grupos, incluindo os licopódios, esfenofilos e progimnospermas, desenvolveram independentemente o "megasporio" – seus esporos eram de dois tamanhos distintos, megásporos maiores e micrósporos menores. Seus gametófitos reduzidos desenvolveram-se a partir de megásporos retidos nos órgãos produtores de esporos (megasporângios) do esporófito, uma condição conhecida como endosporio. As sementes consistem em um megasporângio endospórico cercado por uma ou duas camadas de revestimento (tegumentos). O jovem esporófito se desenvolve dentro da semente, que na germinação se divide para liberá-lo. As primeiras plantas com sementes conhecidas datam do último estágio Devoniano Fameniano. Seguindo a evolução do hábito das sementes, as plantas com sementes diversificaram-se, dando origem a vários grupos hoje extintos, incluindo as samambaias com sementes, bem como as modernas gimnospermas e angiospermas. As gimnospermas produzem "sementes nuas" não totalmente encerrado em um ovário; representantes modernos incluem coníferas, cicadáceas, Ginkgo e Gnetales. As angiospermas produzem sementes encerradas em uma estrutura como um carpelo ou um ovário. Pesquisas em andamento sobre a filogenética molecular de plantas vivas parecem mostrar que as angiospermas são um grupo irmão das gimnospermas.
Fisiologia vegetal

A fisiologia vegetal abrange todas as atividades químicas e físicas internas das plantas associadas à vida. Substâncias químicas obtidas do ar, solo e água formam a base de todo o metabolismo vegetal. A energia da luz solar, captada pela fotossíntese oxigênica e liberada pela respiração celular, é a base de quase toda a vida. Os fotoautotróficos, incluindo todas as plantas verdes, algas e cianobactérias, coletam energia diretamente da luz solar pela fotossíntese. Os heterotróficos, incluindo todos os animais, todos os fungos, todas as plantas completamente parasitas e bactérias não fotossintéticas absorvem moléculas orgânicas produzidas por fotoautotróficos e as respiram ou as utilizam na construção de células e tecidos. A respiração é a oxidação de compostos de carbono, quebrando-os em estruturas mais simples para liberar a energia que eles contêm, essencialmente o oposto da fotossíntese.
As moléculas são movidas dentro das plantas por processos de transporte que operam em uma variedade de escalas espaciais. O transporte subcelular de íons, elétrons e moléculas, como água e enzimas, ocorre através das membranas celulares. Minerais e água são transportados das raízes para outras partes da planta na corrente de transpiração. Difusão, osmose, transporte ativo e fluxo de massa são formas diferentes de ocorrer o transporte. Exemplos de elementos que as plantas precisam transportar são nitrogênio, fósforo, potássio, cálcio, magnésio e enxofre. Nas plantas vasculares, esses elementos são extraídos do solo como íons solúveis pelas raízes e transportados por toda a planta no xilema. A maioria dos elementos necessários para a nutrição das plantas vem da degradação química dos minerais do solo. A sacarose produzida pela fotossíntese é transportada das folhas para outras partes da planta no floema e os hormônios vegetais são transportados por uma variedade de processos.
Hormônios vegetais

2 Com o sol em um ângulo e apenas brilhando em um lado do tiro, auxin se move para o lado oposto e estimula o alongamento celular lá.
3 e 4 O crescimento extra desse lado faz com que o tiro se curve em direção ao sol.
As plantas não são passivas, mas respondem a sinais externos, como luz, toque e lesões, movendo-se ou crescendo em direção ou afastando-se do estímulo, conforme apropriado. Evidência tangível da sensibilidade ao toque é o colapso quase instantâneo dos folhetos de Mimosa pudica, as armadilhas de insetos da armadilha de Vênus e das bexigas, e as polínias das orquídeas.
A hipótese de que o crescimento e desenvolvimento das plantas é coordenado por hormônios vegetais ou reguladores de crescimento vegetal surgiu pela primeira vez no final do século XIX. Darwin experimentou os movimentos dos brotos e raízes das plantas em direção à luz e à gravidade e concluiu: "Não é exagero dizer que a ponta da radícula... age como o cérebro de um dos animais inferiores... dirigindo o vários movimentos". Mais ou menos na mesma época, o papel das auxinas (do grego auxein, para grow) no controle do crescimento das plantas foi delineado pela primeira vez pelo cientista holandês Frits Went. A primeira auxina conhecida, o ácido indol-3-acético (IAA), que promove o crescimento celular, só foi isolada de plantas cerca de 50 anos depois. Este composto medeia as respostas trópicas de brotos e raízes em relação à luz e à gravidade. A descoberta em 1939 de que o calo vegetal poderia ser mantido em cultura contendo IAA, seguida pela observação em 1947 de que ele poderia ser induzido a formar raízes e brotos controlando a concentração de hormônios de crescimento foram passos fundamentais no desenvolvimento da biotecnologia vegetal e da modificação genética..
As citocininas são uma classe de hormônios vegetais nomeados por seu controle da divisão celular (especialmente a citocinese). A citocinina natural zeatina foi descoberta no milho, Zea mays, e é um derivado da purina adenina. A zeatina é produzida nas raízes e transportada para os brotos no xilema, onde promove a divisão celular, o desenvolvimento de brotos e o esverdeamento dos cloroplastos. As giberelinas, como o ácido giberélico, são diterpenos sintetizados a partir da acetil CoA pela via do mevalonato. Eles estão envolvidos na promoção da germinação e quebra de dormência em sementes, na regulação da altura da planta, controlando o alongamento do caule e no controle da floração. O ácido abscísico (ABA) ocorre em todas as plantas terrestres, exceto hepáticas, e é sintetizado a partir de carotenóides nos cloroplastos e outros plastídeos. Inibe a divisão celular, promove a maturação da semente e a dormência e promove o fechamento dos estômatos. Foi assim chamado porque foi originalmente pensado para controlar a abscisão. O etileno é um hormônio gasoso produzido em todos os tecidos vegetais superiores a partir da metionina. Sabe-se agora que é o hormônio que estimula ou regula o amadurecimento e a abscisão dos frutos, e ele, ou o regulador de crescimento sintético ethephon, que é rapidamente metabolizado para produzir etileno, são usados em escala industrial para promover o amadurecimento de algodão, abacaxi e outras culturas climatéricas.
Outra classe de fitohormônios são os jasmonatos, isolados pela primeira vez do óleo de Jasminum grandiflorum, que regulam as respostas a feridas em plantas, desbloqueando a expressão de genes necessários na resposta sistêmica de resistência adquirida ao ataque de patógenos.
Além de ser a principal fonte de energia para as plantas, a luz funciona como um dispositivo de sinalização, fornecendo informações à planta, como a quantidade de luz solar que a planta recebe por dia. Isso pode resultar em mudanças adaptativas em um processo conhecido como fotomorfogênese. Os fitocromos são os fotorreceptores em uma planta que são sensíveis à luz.
Anatomia e morfologia vegetal

A anatomia vegetal é o estudo da estrutura das células e tecidos vegetais, enquanto a morfologia vegetal é o estudo de sua forma externa. Todas as plantas são eucariontes multicelulares, seu DNA armazenado em núcleos. As características das células vegetais que as distinguem das dos animais e dos fungos incluem uma parede celular primária composta pelos polissacarídeos celulose, hemicelulose e pectina, vacúolos maiores do que nas células animais e a presença de plastídeos com funções fotossintéticas e biossintéticas únicas como nas células cloroplastos. Outros plastídeos contêm produtos de armazenamento, como amido (amiloplastos) ou lipídios (elaioplastos). Excepcionalmente, as células estreptófitas e as da ordem de algas verdes Trentepohliales se dividem pela construção de um fragmoplasto como modelo para a construção de uma placa celular no final da divisão celular.
![A diagram of a "typical" eudicot, the most common type of plant (three-fifths of all plant species).[178] However, no plant actually looks exactly like this.](https://en.wikipedia.org/wiki/File:Plant.svg)
Os corpos de plantas vasculares, incluindo clubmosses, samambaias e plantas com sementes (gimnospermas e angiospermas) geralmente têm subsistemas aéreos e subterrâneos. Os brotos consistem em caules com folhas fotossintetizantes verdes e estruturas reprodutivas. As raízes vascularizadas subterrâneas possuem pêlos radiculares em suas pontas e geralmente carecem de clorofila. Plantas não vasculares, hepáticas, antóceros e musgos não produzem raízes vasculares que penetram no solo e a maior parte da planta participa da fotossíntese. A geração de esporófitos não é fotossintética em hepáticas, mas pode ser capaz de contribuir com parte de suas necessidades energéticas pela fotossíntese em musgos e antóceras.
O sistema radicular e o sistema rebento são interdependentes – o sistema radicular geralmente não fotossintético depende do sistema rebento para alimentação, e o sistema rebento geralmente fotossintético depende da água e dos minerais do sistema radicular. As células em cada sistema são capazes de criar células do outro e produzir brotos ou raízes adventícias. Estolões e tubérculos são exemplos de brotos que podem criar raízes. As raízes que se espalham perto da superfície, como as dos salgueiros, podem produzir brotos e, por fim, novas plantas. No caso de um dos sistemas ser perdido, o outro pode recriá-lo. De fato, é possível cultivar uma planta inteira a partir de uma única folha, como é o caso das plantas da seita Streptocarpus. Saintpaulia, ou mesmo uma única célula – que pode se desdiferenciar em um calo (uma massa de células não especializadas) que pode se transformar em uma nova planta. Nas plantas vasculares, o xilema e o floema são os tecidos condutores que transportam recursos entre a parte aérea e as raízes. As raízes são frequentemente adaptadas para armazenar alimentos como açúcares ou amido, como na beterraba e na cenoura.
Os caules fornecem principalmente suporte para as folhas e estruturas reprodutivas, mas podem armazenar água em plantas suculentas como cactos, alimentos como em tubérculos de batata, ou reproduzir vegetativamente como nos estolões de plantas de morango ou no processo de alporquia. As folhas captam a luz do sol e realizam a fotossíntese. Folhas grandes, planas, flexíveis e verdes são chamadas de folhas de folhagem. Gimnospermas, como coníferas, cicas, Ginkgo e gnetófitos são plantas produtoras de sementes com sementes abertas. As angiospermas são plantas produtoras de sementes que produzem flores e têm sementes fechadas. As plantas lenhosas, como as azáleas e os carvalhos, passam por uma fase de crescimento secundário resultando em dois tipos adicionais de tecidos: madeira (xilema secundário) e casca (floema secundário e cortiça). Todas as gimnospermas e muitas angiospermas são plantas lenhosas. Algumas plantas se reproduzem sexuadamente, algumas assexuadamente e algumas por ambos os meios.
Embora a referência às principais categorias morfológicas, como raiz, caule, folha e tricoma, seja útil, deve-se ter em mente que essas categorias estão ligadas por meio de formas intermediárias, de modo que resulta um continuum entre as categorias. Além disso, as estruturas podem ser vistas como processos, ou seja, combinações de processos.
Botânica sistemática

A botânica sistemática faz parte da biologia sistemática, que se preocupa com o alcance e a diversidade dos organismos e suas relações, particularmente conforme determinado por sua história evolutiva. Envolve, ou está relacionado com, classificação biológica, taxonomia científica e filogenética. A classificação biológica é o método pelo qual os botânicos agrupam os organismos em categorias como gêneros ou espécies. A classificação biológica é uma forma de taxonomia científica. A taxonomia moderna está enraizada no trabalho de Carl Linnaeus, que agrupou espécies de acordo com características físicas compartilhadas. Esses agrupamentos foram revisados para se alinhar melhor com o princípio darwiniano de descendência comum – agrupando organismos por ancestralidade em vez de características superficiais. Embora os cientistas nem sempre concordem sobre como classificar os organismos, a filogenética molecular, que usa sequências de DNA como dados, conduziu muitas revisões recentes ao longo das linhas evolutivas e provavelmente continuará a fazê-lo. O sistema de classificação dominante é chamado taxonomia de Lineu. Inclui classificações e nomenclatura binomial. A nomenclatura de organismos botânicos é codificada no Código Internacional de Nomenclatura para Algas, Fungos e Plantas (ICN) e administrado pelo Congresso Botânico Internacional.
O Reino Plantae pertence ao Domínio Eukaryota e é decomposto recursivamente até que cada espécie seja classificada separadamente. A ordem é: Reino; Filo (ou Divisão); Aula; Ordem; Família; Gênero (plural gêneros); Espécies. O nome científico de uma planta representa seu gênero e suas espécies dentro do gênero, resultando em um único nome mundial para cada organismo. Por exemplo, o lírio tigre é Lilium columbianum. Lilium é o gênero e columbianum o epíteto específico. A combinação é o nome da espécie. Ao escrever o nome científico de um organismo, é apropriado colocar a primeira letra do gênero em maiúscula e colocar todos os epítetos específicos em minúsculas. Além disso, o termo inteiro geralmente é colocado em itálico (ou sublinhado quando o itálico não está disponível).
As relações evolutivas e a hereditariedade de um grupo de organismos são chamadas de filogenia. Estudos filogenéticos tentam descobrir filogenias. A abordagem básica é usar semelhanças com base na herança compartilhada para determinar relacionamentos. Por exemplo, espécies de Pereskia são árvores ou arbustos com folhas proeminentes. Obviamente, eles não se assemelham a um típico cacto sem folhas, como um Echinocactus. No entanto, tanto Pereskia quanto Echinocactus têm espinhos produzidos a partir de aréolas (estruturas semelhantes a almofadas altamente especializadas), sugerindo que os dois gêneros estão realmente relacionados.


Julgar relacionamentos com base em personagens compartilhados requer cuidado, pois as plantas podem se assemelhar por meio de uma evolução convergente em que os personagens surgiram de forma independente. Algumas euphorbias têm corpos arredondados e sem folhas, adaptados à conservação de água semelhantes aos dos cactos globulares, mas características como a estrutura de suas flores deixam claro que os dois grupos não estão intimamente relacionados. O método cladístico faz uma abordagem sistemática dos caracteres, distinguindo entre aqueles que não carregam nenhuma informação sobre a história evolutiva compartilhada – como aqueles que evoluíram separadamente em diferentes grupos (homoplasias) ou aqueles que sobraram de ancestrais (plesiomorfias) – e caracteres derivados, que foram transmitidos de inovações em um ancestral compartilhado (apomorfias). Apenas caracteres derivados, como as aréolas produtoras de espinhos de cactos, fornecem evidências de descendência de um ancestral comum. Os resultados das análises cladísticas são expressos como cladogramas: diagramas em forma de árvore mostrando o padrão de ramificação e descendência evolutiva.
A partir da década de 1990, a abordagem predominante para a construção de filogenias para plantas vivas tem sido a filogenética molecular, que usa caracteres moleculares, particularmente sequências de DNA, em vez de caracteres morfológicos como a presença ou ausência de espinhos e aréolas. A diferença é que o próprio código genético é usado para decidir as relações evolutivas, em vez de ser usado indiretamente por meio dos caracteres que ele dá origem. Clive Stace descreve isso como tendo "acesso direto à base genética da evolução". Como um exemplo simples, antes do uso de evidências genéticas, os fungos eram considerados plantas ou estavam mais relacionados a plantas do que a animais. Evidências genéticas sugerem que a verdadeira relação evolutiva de organismos multicelulares é mostrada no cladograma abaixo – os fungos estão mais relacionados aos animais do que às plantas.
| |||||||||||||
Em 1998, o Angiosperm Phylogeny Group publicou uma filogenia para plantas com flores com base na análise de sequências de DNA da maioria das famílias de plantas com flores. Como resultado deste trabalho, muitas questões, como quais famílias representam os primeiros ramos das angiospermas, foram respondidas. Investigar como as espécies de plantas estão relacionadas entre si permite que os botânicos entendam melhor o processo de evolução nas plantas. Apesar do estudo de plantas modelo e do aumento do uso de evidências de DNA, há trabalho e discussão em andamento entre os taxonomistas sobre a melhor forma de classificar as plantas em vários táxons. Os desenvolvimentos tecnológicos, como computadores e microscópios eletrônicos, aumentaram muito o nível de detalhe estudado e a velocidade com que os dados podem ser analisados.
Símbolos
Alguns símbolos são usados atualmente na botânica. Vários outros estão obsoletos; por exemplo, Linnaeus usou símbolos planetários para plantas lenhosas, herbáceas e perenes, e Willd usou ♄ (Saturno) para neutro além de ☿ (Mercury) para hermafroditas. Os seguintes símbolos ainda são usados:
- fêmea
- masculino
- hermafrodite/bisexual
- Reprodução vegetativa (sexual)
- ◊ sexo desconhecido
- anual de solar
 ()) Bienal
()) Bienal ()) perene
()) perene- ous venenoso
- information mais informações
- × híbrido crossbred
- + híbrido enxertado
Contenido relacionado
Par base
Astrobiologia
Kenyanthropus